1. Introduction
Helicobacter pylori is a microaerophilic bacterium and a gastric pathogen that colonizes about 50% of the world’s population.
H. pylori infection causes chronic inflammation and significantly increases the risk of developing duodenal and gastric ulcer diseases and gastric cancer. Chronic
H. pylori infection is now considered to be the strongest risk factor for gastric cancer (Wroblewski et al.,
2010). Recent reports have suggested that exacerbated DNA damage of gastric cells results from inflammation-related oxidative stress induced by reactive oxygen species (ROS)
in vivo (Coussens and Werb,
2002).
During the process of colonizing and infiltrating the host cells by
H. pylori, innate and adaptive inflammatory responses are activated. The activated neutrophils of the host can produce ROS and reactive nitrogen species (RNS), and it has been reported that
H. pylori-induced ROS and RNS production in gastric epithelial cells might affect signal transduction and damage DNA in gastric cells, resulting in gastric carcinogenesis (Handa et al.,
2010). The inflammatory host response can not only lead to gastric cancer, but also threaten the survival of
H. pylori. The suppressing effect of oxidative stress species has also been reported
in vitro (Qu et al.,
2011). Nevertheless,
H. pylori successfully survives these conditions and persistently colonizes the gastric mucosa. Therefore, mechanisms for detoxification of redox compounds and repair of damaged cell components in
H. pylori are particularly important in understanding
H. pylori pathogenesis and persistence (Wang et al.,
2006).
The antioxidant systems in
H. pylori appear to be essential for its survival in the host. Studies have indicated that these systems have many components. Thioredoxin-1 (Trx1) is one of the antioxidants of
H. pylori (Comtois et al.,
2003).
H. pylori Trx1 is similar to a eukaryotic peroxidase reduction system. Trx1 contains a redox-active site, Cys-Gly-Pro-Cys, and serves as a ubiquitous enzyme that catalyzes the reduction of disulfide bonds (Alamuri and Maier,
2006). Using the purified
H. pylori Trx1 and alkyl hydroperoxide reductase (AhpC), Baker et al. (
2001) found that Trx1 can transfer electrons to AhpC. AhpC is an enzyme that reduces peroxides and can protect bacterial and human cells against reactive nitrogen intermediates (Chen et al.,
1998).
H. pylori Trx1 has also been reported as an arginase (RocF) chaperone. Trx1 and arginase equip
H. pylori with a “redox guardian” to overcome oxidative and nitrosative stress (McGee et al.,
2006). The functions and relationships of these antioxidants have been reported. However, there is little direct evidence showing the expression of these antioxidants in clinical isolates and their associations with clinical gastric diseases.
In this study, we investigated
Trx1,
RocF, and
AhpC expression in
H. pylori strains isolated from tissues exhibiting gastritis, peptic ulcer, and gastric cancer. The relationships among the expression of these antioxidants in
H. pylori were also assessed using Pearson’s correlation test.
2. Materials and methods
2.1. Patient samples
Gastric biopsies were obtained from a total of 61 patients with gastric cancer, peptic ulcer, and gastritis, who had undergone gastroscopic examination at Peking University Third Hospital (Beijing, China) from 2010 to 2012. All collected tissues had positive results from a rapid urease test, which were confirmed by Warthin-Starry staining. Written informed consent with a signature was obtained from all patients. All tissues were assessed by hematoxylin-eosin (HE) staining. Diagnoses of all the samples were confirmed histologically by two independent pathologists.
2.2. Culturing of bacterial strains
Two antral biopsies used for
H. pylori culture were collected in tubes containing brucella broth. After homogenizing immediately, the samples were plated on 5% (v/v) sheep blood agar plates containing amphotericin B 4 μg/ml, trimethoprim 4 μg/ml, and vancomycin 4 μg/ml (Life Tech, Carlsbad, CA, USA). The plates were incubated for 5 to 7 d at 37 °C under microaerophilic conditions using a microaerobic pack (Mitsubishi Gas Chemical Co., Inc., Japan).
H. pylori cultures were examined using urease tests and Gram staining. Oxidase tests and catalase tests were used to ensure that the cultures were not contaminated. Bacterial culture was successful in samples from 40 out of 61 infected patients. The 40 samples were isolated from 14 tissues displaying gastritis (including 9 superficial gastritis tissues and 5 atrophic gastritis tissues), 13 peptic ulcer tissues (including 8 gastric ulcer tissues and 5 duodenal ulcer tissues), and 13 gastric cancer tissues (including 2 tissues at an early stage and 11 tissues at an advanced stage; 6 intestinal type tissues, 4 diffuse type tissues, and 3 mixed type tissues).
2.3. RNA isolation
RNA extraction from
H. pylori was performed as described previously (Shi et al.,
2013). Briefly,
H. pylori strains were resuscitated on blood agar plates and harvested to precooled sterile phosphate buffered saline (PBS). After washing with PBS twice, total RNA was isolated from
H. pylori strains using TRIzol (Invitrogen) following the manufacturer’s instructions. RNAs were treated with RNase-free DNase I (Fermentas). The quality of RNA was determined by agarose gel electrophoresis and the quantity by spectrophotometry at 260 nm. RNA was stored at −80 °C until needed.
2.4. Reverse transcription and real-time polymerase chain reaction (PCR)
Reverse transcription and real-time PCR procedures were performed as described previously (Shi et al.,
2013; Zhang et al.,
2013). Total RNA (2 μg) was reverse-transcribed using the RevertAid First Strand cDNA Synthesis Kit (Fermentas). Real-time PCR was performed using SYBR Green on a Lightcycler 480II Real-Time PCR Detection System (Roche, Indianapolis, IN, USA). All results were normalized to 16S rRNA amplification. Each reaction was carried out in technical triplicates. Primers were as follows:
H. pylori Trx1: 5′-GGGGTTGCGTTAGTGGATTTTTG-3′ (forward primer) and 5′-GACGACTTCGCCATCTTTTGTGA-3′ (reverse primer);
H. pylori RocF: 5′-TTTACCTTAGCCTGGATTTAGACA-3′ (forward primer) and 5′-GTTGTATTCGGTTACTTCAAGTGC-3′ (reverse primer);
H. pylori AhpC: 5′-CCCTACAGAAATCATTGCGTTTG-3′ (forward primer) and 5′-GGAAAGACACTTGACCAATACCG-3′ (reverse primer); 16S rRNA: 5′-CCGCCTACGCGCTCTTTAC-3′ (forward primer) and 5′-CTAACGAATAAGCACCGGCTAAC-3′ (reverse primer). The relative expression of the target genes was calculated based on cycle threshold (
C
T) measurements. The ∆
C
T values of each sample were calculated as
C
T,target gene−
C
T,16S rRNA (Shen et al.,
2010).
2.5. Statistical analysis
A one-way analysis of variance (ANOVA) test followed by a Student-Newman-Keuls (SNK) test was used to analyze the significant differences among multiple sample sets. Pearson’s correlation test was used to assess the relationships between multiple continuous variables. Data were presented as mean±standard deviation (SD) of three independent experiments. All statistical analyses were performed using SPSS 13.0 software (SPSS Inc., Chicago, IL, USA).
P-values less than 0.05 were considered statistically significant. GraphPad Prism 5 was used to create the artwork.
3. Results
3.1. Expression of Trx1, RocF, and AhpC in H. pylori strains isolated from tissues displaying gastric cancer, peptic ulcer, and gastritis
H. pylori was isolated from 14 gastritis samples, 13 peptic ulcer samples, and 13 gastric cancer samples, and the expression of
Trx1,
RocF, and
AhpC was evaluated.
Trx1 and
AhpC expression was successfully measured in all 40 strains. For the expression of
RocF, 33 strains out of 40 were successfully evaluated. The mRNA expression level of
Trx1 in
H. pylori isolated from the gastric cancer and peptic ulcer tissues was higher (
P<0.05) than that in gastritis tissues (Fig.
1a). No significant difference in
Trx1 expression was found between the gastric cancer and peptic ulcer samples. The mRNA expression level of
RocF in
H. pylori isolated from the gastric cancer samples was significantly higher (
P<0.05) than the level observed for tissues exhibiting gastritis and peptic ulcer (Fig.
1b). However, no significant difference in
RocF expression was found between the gastritis and peptic ulcer samples. The mRNA expression levels of
AhpC in
H. pylori isolated from the gastritis, peptic ulcer, and gastric cancer samples showed no significant differences (Fig.
1c).
Fig.1
Trx1 (a), RocF (b), and AhpC (c) expression of H. pylori in gastric diseases (gastritis, peptic ulcer, and gastric cancer) evaluated by real-time PCR
The mRNA was extracted from H. pylori strains isolated from 14 gastritis samples, 13 peptic ulcer samples, and 13 gastric cancer samples. The Trx1 (a), RocF (b), and AhpC (c) expression of H. pylori was evaluated in these tissues by real-time PCR and normalized to the control gene 16S rRNA. The ΔC
T value of each sample was calculated as C
T,target gene−C
T,16S rRNA. We successfully detected Trx1 and AhpC in all 40 strains and RocF in 33 strains out of 40. * When compared with tissues displaying gastritis, P<0.05 was considered statistically significant. # When compared with peptic ulcer samples, P<0.05 was considered statistically significant
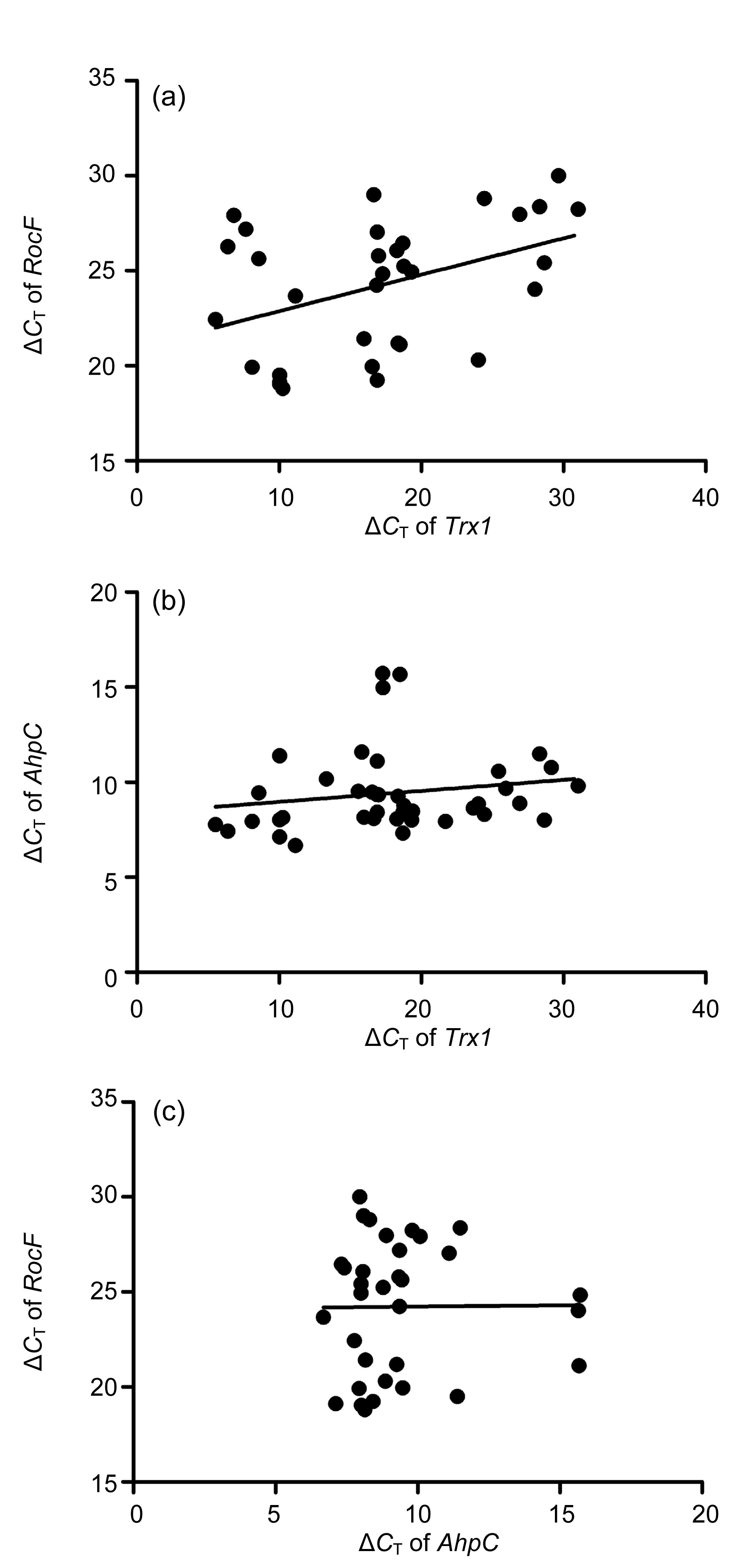
3.2. Correlation among the expression of RocF, Trx1, and AhpC in H. pylori strains
The relationships between
Trx1,
RocF, and
AhpC expression in
H. pylori strains were assessed using Pearson’s correlation test.
Trx1 expression had a positive, linear correlation with
RocF expression (
r=0.411,
P<0.05; Fig.
2a).
Trx1 expression had a low positive correlation with
AhpC expression (
r=0.178), but did not present a linear trend (
P>0.05; Fig.
2b), However, there was no significant correlation (
r=0.009;
P>0.05) between the expression of
RocF and
AhpC (Fig.
2c).
Fig.2
Correlation analysis of Trx1, RocF, and AhpC expression in clinical H. pylori isolates
(a) The expression of Trx1 in H. pylori strains had a positive, linear correlation with the expression of RocF; (b) The expression of Trx1 in H. pylori strains had a weakly positive correlation with the expression of AhpC, but there was no linear trend; (c) The expression of AhpC in H. pylori strains was not significantly correlated with the expression of RocF
4. Discussion
It is well known that
H. pylori infection induces progressive inflammatory changes in the gastric mucosa and increases the risk of peptic ulcer disease and gastric cancer. The antioxidant systems in
H. pylori were expected to be relevant to bacterial-associated, and especially inflammation-associated, infectious diseases (Wang et al.,
2006). In a previous study, we investigated the comparative proteome of clinical
H. pylori strains by two-dimensional gel electrophoresis. We reported differential proteins of
H. pylori strains isolated from gastritis and gastric cancer, including antioxidants (Zhang et al.,
2011). In this study, we investigated the expression of three essential antioxidants of
H. pylori in clinical isolates to show the relationship between the expression of these antioxidants and different gastric diseases. Correlations between the expression of these antioxidants were also assessed.
Our results showed that
Trx1 expression of
H. pylori in clinical gastric cancer and peptic ulcer tissues was higher than that in gastritis tissues.
RocF expression of
H. pylori in gastric cancer tissues was higher than that in tissues exhibiting peptic ulcer and gastritis. Furthermore, Pearson’s correlation test showed that
Trx1 and
RocF expression had a positive, linear correlation. Trx1 is a key protein in many crucial cellular functions, including oxidative stress management (Ritz and Beckwith,
2001). Studies have shown that the expression of intracellular
H. pylori Trx1 is altered in response to a variety of applied extracellular stresses, suggesting that Trx1 behaves as a stress response element in
H. pylori (Windle et al.,
2000). Moreover, Trx1 of
H. pylori is considered to be an arginase chaperone because of its active site dithiol/disulfide (Berndt et al.,
2008). The RocF protein arginase hydrolyzes arginine to ornithine and urea, and urease hydrolyzes urea to carbon dioxide and ammonium, which can neutralize acid. Several reports have shown that the enzymatic pathway of arginase used by
H. pylori for the production of ornithine and urea can also serve as a mechanism responsible for the lack of protective effect of the immune response and the chronicity of
H. pylori infection by inhibiting T cell proliferation and by down-regulating eukaryotic NO production (Gobert et al.,
2001; Zabaleta et al.,
2004). There is a critical disulphide bond for the stability of arginase, which is modulated at the post-translational level by Trx1. Trx1 can convert denatured or suboptimally folded arginase into an optimal three-dimensional catalytically active structure (McGee et al.,
2006). In this study, we investigated the relationship between
Trx1 and
RocF expression in clinical isolates of
H. pylori, based on the supposition that the expression of
Trx1 and
RocF might be correlated and might relate to gastric carcinogenesis.
In this study, we isolated 40 strains from gastric tissues. It has been reported that
H. pylori RocF in clinical isolates has genetic microheterogeneity and phenotypic variation (Hovey et al.,
2007). To avoid genetic variation in
RocF affecting our PCR performance, we designed real-time PCR primers for
RocF in its conserved region suggested by gene sequencing (Hovey et al.,
2007). For the expression of
RocF, only 33 strains out of 40 were successfully evaluated, probably because of the generally low expression of
RocF shown by the PCR results in this work.
In addition to
RocF, we investigated the expression of
AhpC, one of the most abundant proteins expressed in
H. pylori and an antioxidant enzyme whose function is essential for reducing different peroxides (Baker et al.,
2001). Huang and Chiou (
2011) reported the upregulation of
H. pylori AhpC after treatment with hydrogen peroxide, which suggested that AhpC might play a part in protecting organisms from damage by ROS.
AhpC mutants are hypersensitive to oxygen and organic peroxides and are restricted to atmospheres of less than 2% oxygen for growth (Olczak et al.,
2002). AhpC might assume an important role in combating exogenous peroxides arising from lifelong chronic inflammation (Croxen et al.,
2007). Further study using 2-dimensional gel electrophoresis (2-DE) showed that AhpC was expressed in higher amounts in
H. pylori strains isolated from three patients with gastric cancer than in strains from three patients with gastritis (Huang et al.,
2011). In this study, we assessed the expression of
AhpC using real-time PCR in 40 clinical strains from patients, including 13 displaying gastric cancer, 13 displaying peptic ulcer, and 14 displaying gastritis. However, our data did not show any differences in
AhpC expression in
H. pylori isolated from gastric cancer, peptic ulcer, and gastritis tissues. Although it has been reported that purified Trx1 can transfer electrons to AhpC (Baker et al.,
2001), we did not find any relationship between the expression of
AhpC and
Trx1 in clinical
H. pylori isolates. Some previous studies indicated that AhpC was essential for both
in vitro growth and gastric colonization (Chalker et al.,
2001). However, Croxen et al. (
2007) showed that high levels of AhpC were not required for
in vitro growth or for primary gastric colonization. The functions of AhpC remain unclear and are worthy of further study.
In conclusion, our findings demonstrate that the expression of
H. pylori Trx1 and
RocF is higher in gastric cancer tissues than in gastritis tissues.
H. pylori infection can induce inflammatory responses followed by ROS/RNS and DNA damage in tissues. Nevertheless,
H. pylori successfully survives these conditions by means of antioxidant enzymes, chronically infects gastric epithelium, and induces severe gastric outcomes. This study lends support to the supposition that the antioxidant enzymes in
H. pylori may be related to the development of gastric cancer from
H. pylori infection.
Trx1 and
RocF of
H. pylori may prove to be prognostic for monitoring varied clinical manifestations of gastrointestinal patients infected with
H. pylori. Further studies should performed to confirm the relationship between
H. pylori Trx1 and RocF, and to gain more insight into the functions and mechanisms of action of
H. pylori Trx1 and RocF on gastric mucosa, using cell experiments and animal models. Research related to clinical patients and diseases should facilitate a better understanding of the involvement of
H. pylori antioxidants in pathogenic mechanisms.
Acknowledgements
We wish to thank Feng-min LU of Peking University (China) for providing technological help.
* Project supported by the National Natural Science Foundation of China (No. 81270475)Compliance with ethics guidelines Yan-yan SHI, Mo CHEN, Yue-xia ZHANG, Jing ZHANG, and Shi-gang DING declare that they have no conflict of interest.References
[1] Alamuri, P., Maier, R.J., 2006. Methionine sulfoxide reductase in
Helicobacter pylori: interaction with methionine-rich proteins and stress-induced expression.
J Bacteriol, 188(16):5839-5850.


[2] Baker, L.M., Raudonikiene, A., Hoffman, P.S., 2001. Essential thioredoxin-dependent peroxiredoxin system from
Helicobacter pylori: genetic and kinetic characterization.
J Bacteriol, 183(6):1961-1973.
[3] Berndt, C., Lillig, C.H., Holmgren, A., 2008. Thioredoxins and glutaredoxins as facilitators of protein folding.
Biochim Biophys Acta, 1783(4):641-650.
[4] Chalker, A.F., Minehart, H.W., Hughes, N.J., 2001. Systematic identification of selective essential genes in
Helicobacter pylori by genome prioritization and allelic replacement mutagenesis.
J Bacteriol, 183(4):1259-1268.
[5] Chen, L., Xie, Q.W., Nathan, C., 1998. Alkyl hydroperoxide reductase subunit C (AhpC) protects bacterial and human cells against reactive nitrogen intermediates.
Mol Cell, 1(6):795-805.
[6] Comtois, S.L., Gidley, M.D., Kelly, D.J., 2003. Role of the thioredoxin system and the thiol-peroxidases Tpx and Bcp in mediating resistance to oxidative and nitrosative stress in
Helicobacter pylori
.
Microbiology, 149(1):121-129.
[7] Coussens, L.M., Werb, Z., 2002. Inflammation and cancer.
Nature, 420(6917):860-867.
[8] Croxen, M.A., Ernst, P.B., Hoffman, P.S., 2007. Antisense RNA modulation of alkyl hydroperoxide reductase levels in
Helicobacter pylori correlates with organic peroxide toxicity but not infectivity.
J Bacteriol, 189(9):3359-3368.
[9] Gobert, A.P., McGee, D.J., Akhtar, M., 2001.
Helicobacter pylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterial survival.
PNAS, 98(24):13844-13849.
[10] Handa, O., Naito, Y., Yoshikawa, T., 2010.
Helicobacter pylori: a ROS-inducing bacterial species in the stomach.
Inflamm Res, 59(12):997-1003.
[11] Hovey, J.G., Watson, E.L., Langford, M.L., 2007. Genetic microheterogeneity and phenotypic variation of
Helicobacter pylori arginase in clinical isolates.
BMC Microbiol, 7(1):26
[12] Huang, C.H., Chuang, M.H., Lo, W.L., 2011. Alkylhydroperoxide reductase of
Helicobacter pylori as a biomarker for gastric patients with different pathological manifestations.
Biochimie, 93(7):1115-1123.
[13] McGee, D.J., Kumar, S., Viator, R.J., 2006.
Helicobacter pylori thioredoxin is an arginase chaperone and guardian against oxidative and nitrosative stresses.
J Biol Chem, 281(6):3290-3296.
[14] Olczak, A.A., Olson, J.W., Maier, R.J., 2002. Oxidative-stress resistance mutants of
Helicobacter pylori
.
J Bacteriol, 184(12):3186-3193.
[15] Qu, W., Zhou, Y., Sun, Y., 2011. Identification of
S-nitrosylation of proteins of
Helicobacter pylori in response to nitric oxide stress.
J Microbiol, 49(2):251-256.
[16] Ritz, D., Beckwith, J., 2001. Roles of thiol-redox pathways in bacteria.
Ann Rev Microbiol, 55(1):21-48.
[17] Shen, R., Pan, S., Qi, S., 2010. Epigenetic repression of microRNA-129-2 leads to overexpression of SOX4 in gastric cancer.
Biochem Biophys Res Commun, 394(4):1047-1052.
[18] Shi, Y., Liu, L., Zhang, T., 2013. The involvement of
Helicobacter pylori thioredoxin-1 in gastric carcinogenesis.
J Med Microbiol, 62(Pt 8):1226-1234.
[19] Wang, G., Alamuri, P., Maier, R.J., 2006. The diverse antioxidant systems of
Helicobacter pylori
.
Mol Microbiol, 61(4):847-860.
[20] Windle, H.J., Fox, A., NıEidhin, D., 2000. The thioredoxin system of
Helicobacter pylori
.
J Biol Chem, 275(7):5081-5089.
[21] Wroblewski, L.E., Peek, R.M., Wilson, K.T., 2010.
Helicobacter pylori and gastric cancer: factors that modulate disease risk.
Clin Microbiol Rev, 23(4):713-739.
[22] Zabaleta, J., McGee, D.J., Zea, A.H., 2004.
Helicobacter pylori arginase inhibits T cell proliferation and reduces the expression of the TCR ζ-chain (CD3ζ).
J Immunol, 173(1):586-593.
[23] Zhang, H.H., Cai, A.Z., Wei, X.M., 2013. Characterization of cancer stem-like cells in the side population cells of human gastric cancer cell line MKN-45.
J Zhejiang Univ-Sci B (Biomed & Biotechnol), 14(3):216-223.
[24] Zhang, Y.N., Ding, S.G., Huang, L.H., 2011. Comparative proteome analysis of
Helicobacter pylori clinical strains by two-dimensional gel electrophoresis.
J Zhejiang Univ Sci B (Biomed & Biotechnol), 12(10):820-827.


Open peer comments: Debate/Discuss/Question/Opinion
<1>