1. Introduction
Angiotensin-I-converting enzyme (ACE; EC 3.4. 15.1) is an important enzyme in the renin-angiotensin system, which plays a pivotal role in the regulation of blood pressure. The enzyme raises blood pressure by catalyzing the conversion of inactive angiotensin-I to angiotensin-II, a powerful endogenous vasoconstrictor. It is also involved in the release of a sodium-retaining steroid and aldosterone from the adrenal cortex, which has a tendency to stimulate blood vessels to shrink and increase blood pressure (Ondetti and Cushman,
1982). Increased serum ACE levels are associated with hypertension and other diseases, such as congestive heart failure, left ventricular hypertrophy, and acute myocardial infarction (Roškar et al.,
2009). Captopril, lisinopril, enalapril, and fosinopril are chemically synthesized pharmaceuticals commonly used in the treatment of hypertension (Atkinson et al.,
1980). However, these synthetic ACE inhibitors may cause side effects, such as coughing, taste disturbances, skin rashes, and angioneurotic edema. In addition, their use may lead to the aldosterone escape phenomenon and reduce the efficacy of ACE inhibitors after a long-term administration (Chen et al.,
2012).
Many studies on ACE inhibitors from food sources have been undertaken since the first ACE inhibitor was isolated from snake venom (Ferreira,
1965). It was first reported that peptides derived from digests of gelatin exhibited ACE inhibitory activity (Oshima et al.,
1979). Subsequently, ACE inhibitory peptides have been prepared from many other food sources such as soy, milk, fish muscle, and cereals (Ichimura et al.,
2003; Zhang et al.,
2006; Costa et al.,
2007; Tovar-Pérez et al.,
2009). ACE inhibitory peptides may have broad application in the treatment of hypertension (Li et al.,
2004), due to their potential physiological and pharmacological effects.
During the past decade, food scientists have obtained ACE inhibitory peptides from different food sources by a variety of methods, and have developed a complete system for their preparation, separation, and purification (Jang and Lee,
2005; Chen et al.,
2007; Lee et al.,
2009; Jiang et al.,
2010). Heat treatment is one of the most common processing steps in the food industry, and is commonly used for food safety, drying, or stabilization purposes. It is unclear whether ACE inhibitory peptides would maintain their inhibitory activities, change their chemical composition, produce browning, or cause toxicity during heat treatment of food during processing in the presence of acid or alkali. In this study, we tested ACE inhibitory peptides derived from bovine casein that had been validated as having strong antihypertensive activity (Jiang et al.,
2010). The objective of this study was to evaluate the processing stability and cell viability of bovine casein-derived ACE inhibitory peptides under different heat treatments. The results might be useful in evaluating the use of ACE inhibitory peptides from bovine casein as active ingredients in functional foods with potential antihypertensive effects.
2. Materials and methods
2.1. Materials
Bovine casein was prepared from bovine milk using an isoelectric precipitation method developed by the Key Laboratory of Dairy Science, Northeast Agricultural University, Harbin, China. AS1.398 neutral protease (from
Bacillus subtilis; activity 50 000 U/g) was purchased from the Wuxi Enzyme Preparation Company (Wuxi, China). Caco-2 cells (TCHu146), a human colon adenocarcinoma cell line, were purchased from the Shanghai Institutes for Biological Sciences of the Chinese Academy of Sciences. Human umbilical vein endothelial ECV-304 cell lines were provided by Dr. Shu-jing WANG from Harbin University of Commerce, China. Hippuryl-
L-hisstidyl-
L-leucine (Hip-His-Leu), ACE, hippuric acid (HA),
o-phthaldialdehyde (OPA),
L-leucine, β-mercaptoethanol, 3-(4,5-dimethythiazol-2-yl)-2,5-diphenyltetr-azolium bromide (MTT), and sodium dodecyl sulphate (SDS) were purchased from Sigma-Aldrich Co. (USA). All other reagents were of analytical grade.
2.2. Preparation of bovine casein-derived ACE inhibitory peptides
ACE inhibitory peptides were prepared by the method of Jiang et al. (
2010). Casein (75 g/L) was dissolved in pH 7.0, 0.04 mol/L phosphate buffer. Hydrolysis was initiated by addition of AS1.398 neutral protease at 45 °C and the enzyme concentration was fixed at 5% (w/w protein). The pH value was kept at 7.0 by continuous addition of 1 mol/L NaOH. After the hydrolysis reaction, the protein hydrolysates were heated at 95 °C for 20 min to inactivate this enzyme and then centrifuged at 4000
g for 20 min at 4 °C. Subsequently, the supernatants were ultrafiltrated through a 3-kDa molecular weight cut-off membrane (Millipore Co., USA) at 35 °C under a pressure of 3 MPa. The resulting permeates were collected and lyophilised to obtain the bovine casein-derived ACE inhibitory peptides.
2.3. Stability of bovine casein-derived ACE inhibitory peptides
To investigate the effect of temperature, samples of bovine casein-derived ACE inhibitory peptide solutions (80 g/L) were adjusted to pH 7.0 and heated at 40, 60, 80, 90, 100, 110, or 120 °C for 2 h. To investigate the effect of pH, peptide solutions were heated at 100 °C at pH values of 1.0, 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, 11.0, or 12.0 for 1 h. To investigate the effect of the duration of heat treatment, peptide solutions were adjusted to pH 7.0 and incubated at 100 °C for 0.5, 1.0, 2.0, 3.0, or 4.0 h. Unheated peptides were used as controls. In all the experiments, each treatment was carried out using triplicate samples.
2.4. Determination of ACE inhibitory activity
ACE inhibitory activity was measured by high-performance liquid chromatography (HPLC), using a modification of the method of Cushman and Cheung (
1971). For each assay, a mixture containing 120 μl of 5.0 mmol/L Hip-His-Leu dissolved in 50 mmol/L sodium borate buffer (pH 8.3) and 20 μl of peptide solution was preincubated at 37 °C for 5 min. ACE solution (100 U/L) of 10 μl was then added to the mixture and incubated at 37 °C for 1 h. Thereafter, the reaction was terminated by the addition of 150 μl of 1 mol/L HCl. Reaction samples were filtered through 0.45 μm filters and 50 μl were injected. Hippuric acid liberated by ACE was determined by reverse-phase HPLC (RP-HPLC) onto a Yilite SinoChrom ODS-BP C18 column (250 mm×4.6 mm i.d., 5 μm, Dalian, China) at 30 °C. A linear gradient from 20% to 70% acetonitrile containing 0.1% trifluoroacetic acid (TFA) was applied in 10 min, and then from 70% to 0% acetonitrile containing 0.1% TFA was reached in 2 min. Flow rate was 1.0 ml/min and elution was monitored at 228 nm. In all experiments, each treatment was carried out with triplicate samples. The percentage of ACE inhibitory activity was calculated as (
A
0−
A
1)×100%/
A
0, where
A
0 represents the absorbance of hippuric acid without the sample and
A
1 represents the absorbance of hippuric acid with the sample.
2.5. Determination of free amino groups
The content of free amino groups was determined by the OPA method with some modifications (Church et al.,
1983). The OPA reagent was prepared to a final volume of 100 ml with deionized water, and consisted of 5.0 ml of 200 mg/L SDS, 80 mg of OPA in 2 ml methanol, 200 μl of β-mercaptoethanol, and 50 ml of 100 mmol/L sodium borate buffer (pH 9.5). Then, 3 ml of OPA reagent was mixed with 100 μl of 2 mg/L ACE inhibitory peptide solution. After vortexing and a minimum 5 min delay in darkness at room temperature, the absorbance was recorded at 340 nm by an ultraviolet (UV) spectrophotometer (UV-2401PC, Shimadzu, Japan). Calibration curves were obtained by using 0–0.55 g/L leucine as a standard, and absorbance readings were converted into free amino group contents. The changes in free amino groups were expressed as relative concentrations (%) in comparison with the content of such groups in unheated ACE inhibitory peptides.
2.6. Determination of color
Colormetric measurements of ACE inhibitory peptide samples were made using an SE-2000 color difference meter (Nippon Denshoku Industries Co., Ltd., Japan). The color difference meter provides the values of three color components: the
L
* value represents black and white components, the
a
* value represents red to green components, and the
b
* value represents yellow to blue components. Samples of 2 ml with a concentration of 20 mg/ml were measured in a quartz cuvette. All samples of each treatment were read six times.
2.7. Cell culture and cell viability assay in vitro
The cell viability of ACE inhibitory peptides was determined by MTT assay against Caco-2 and ECV-304 cell lines. The two kinds of cells were cultured in Dulbecco’s modification of Eagle’s medium (DMEM; GIBCO, USA) containing 100 U/ml penicillin, 100 g/ml streptomycin, and 100 ml/L fetal bovine serum (FBS), and maintained at 37 °C in a humidified atmosphere with 5% CO
2. Cells in logarithmic growth were chosen and cultured in the 96-well plates at a density of 2.5×10
3 cells per well. When confluence was up to 60%–70%, the culture solutions were removed and freshly prepared DMEM, with different concentrations of ACE inhibitory peptides, was added to the plates. After incubation for 48 h, 20 μl of 5 mg/ml MTT solutions was added to each well and cultivation continued for 4 h. Afterwards, the MTT solutions were carefully removed and 150 μl of dimethylsulphoxide (DMSO) was added to each well to dissolve the formazan crystals. The optical density was assayed with a microplate reader (Model 680, Bio-Rad Laboratories, Hercules, USA) at 490 nm. The cell viability was calculated as
A
sample/
A
control×100%. All the MTT assays were repeated at least six times.
2.8. Statistical analysis
Three independent experiments were conducted to test the stability and cytotoxicity of ACE inhibitory peptides derived from bovine casein. One-way analysis of variance (ANOVA) and independent samples
t-tests were conducted to determine the significance of the main effects, using the statistical analysis system SPSS 13.0 for windows (SPSS Inc., Chicago, IL). Data are expressed as mean±standard deviation (SD). Significant differences (
P<0.05) among means were identified using Duncan’s multiple range tests and independent sample
t-tests.
3. Results and discussion
3.1. Effects of heat treatment on the ACE inhibitory activity of bovine casein-derived peptides
Bovine casein-derived peptides can be delivered in the form of functional ingredients, or incorporated into other food products. During thermal processing, it is important to know whether their bioactivities can be maintained. The effects of different pH values (1.0 to 12.0) on the ACE inhibitory activity of bovine casein-derived peptides at 100 °C after 1 h of heat treatment are shown in Fig.
1a. There were no significant differences between the ACE inhibitory activity of the heated peptides in the pH range of 1.0 to 8.0 and that of unheated peptides (
P>0.05). This indicates that bovine casein-derived peptides can resist the acid or weak alkaline conditions of food processing. Hwang (
2010) reported that tuna cooking juice-derived ACE inhibitory peptides had some resistance to the influence of pH. Peptide mapping using HPLC showed slight differences before and after treatments at pH 2.0 and 10.0. Similar results demonstrated that hendeca-peptide isolated from the pepsin hydrolysate of algae protein waste completely retained its ACE inhibitory activity in the pH range of 2.0 to 10.0 (Sheih et al.,
2009). However, the ACE inhibitory activities of peptides noticeably decreased within the pH range of 9.0 to 12.0 (
P<0.05). When the pH value reached 12.0, the peptides lost 20.16% ACE inhibitory activity, compared with the unheated controls. These results indicated that strong alkaline conditions could considerably reduce the ACE inhibitory activities of bovine casein-derived peptides. This is probably due to the degradation or hydrolysis of some peptides into inactive fragments under strong alkaline heating.
Fig.1
Changes in inhibitory activity of ACE inhibitory peptides
(a) At different pH levels (1.0–12.0), 100 °C for 1 h of heat treatment; (b) At different temperatures (40–120 °C), pH 7.0 for 2 h; (c) For different heating time (0–4 h) at 100 °C and pH 7.0. C denotes the ACE inhibitory peptides without heat treatment. Values are expressed as mean±SD, n=3. Different letters above the bars are significantly different (P<0.05)
Heat treatment is the most important method in food processing. It causes protein denaturation and aggregation during temperature changes from 60 to 90 °C (Boye et al.,
1997; Korhonen et al.,
1998). Effects of different temperatures (40 to 120 °C) on the ACE inhibitory activity of bovine casein-derived peptides at pH 7.0 after 2 h of heat treatment are shown in Fig.
1b. There were no significant differences in ACE inhibitory activity between bovine casein-derived peptides heated to 40–100 °C (
P>0.05) and unheated peptides. Similarly, Jang et al. (
2007) found that the activities of peptides DFHINQ and GFHI from beef sarcoplasmic protein extracts showed no significant change after heating at 70, 80, 90, or 100 °C for 20 min. In addition, the activity of soy protein-derived ACE inhibitory peptides retained its activity under various temperature treatments (Wu and Ding,
2002). However, some severe heat treatments of ACE inhibitory peptides, especially in preserve sterilization, might result in a decrease in activity due to the loss of available peptides compared with other more mild treatments (Hwang,
2010). In this study, when bovine casein-derived peptides were heated at 110 and 120 °C, the ACE inhibitory activities of peptides decreased by about 6.55% and 14.14%, respectively (
P<0.05), compared to that of unheated peptides. Therefore, temperature may be a crucial factor that may lead to the loss of ACE inhibitory activity of bovine casein-derived peptides. Nevertheless, the mechanism of the structure-activity relationship of ACE inhibitory peptides is still unclear. The C-terminal sequence is known to be the factor that most strongly influences the binding to ACE. In addition, the inhibitory potency may be augmented by the presence of the positive charge of Lys and Arg as the C-terminal residue. Thus, high heating may potentially alter the C-terminal sequence so that the ACE inhibitory activities of peptides are greatly affected (López-Fandino et al.,
2006).
Heating time is also a major factor that may affect the ACE inhibitory activity of casein-derived peptides. The effects of different heating time (0.5 to 4.0 h) on the ACE inhibitory activity of bovine casein-derived peptides at 100 °C and pH 7.0 are shown in Fig.
1c. Compared to the unheated peptides, there was no statistically significant loss of ACE inhibitory activity in the bovine casein-derived peptides under heat treatments of 0.5 to 3.0 h (
P>0.05), while heat treatment for 4.0 h slightly decreased activity (
P<0.05). The results showed that bovine casein-derived peptides could maintain ACE inhibitory activity for 3 h at pH 7.0 and at 100 °C. In contrast, Akıllıoğlu and Karakaya (
2009) demonstrated that ACE inhibitory activity of legume species increased after heat treatment for 50 min. Therefore, heating time may affect the biological activity of ACE inhibitory peptides.
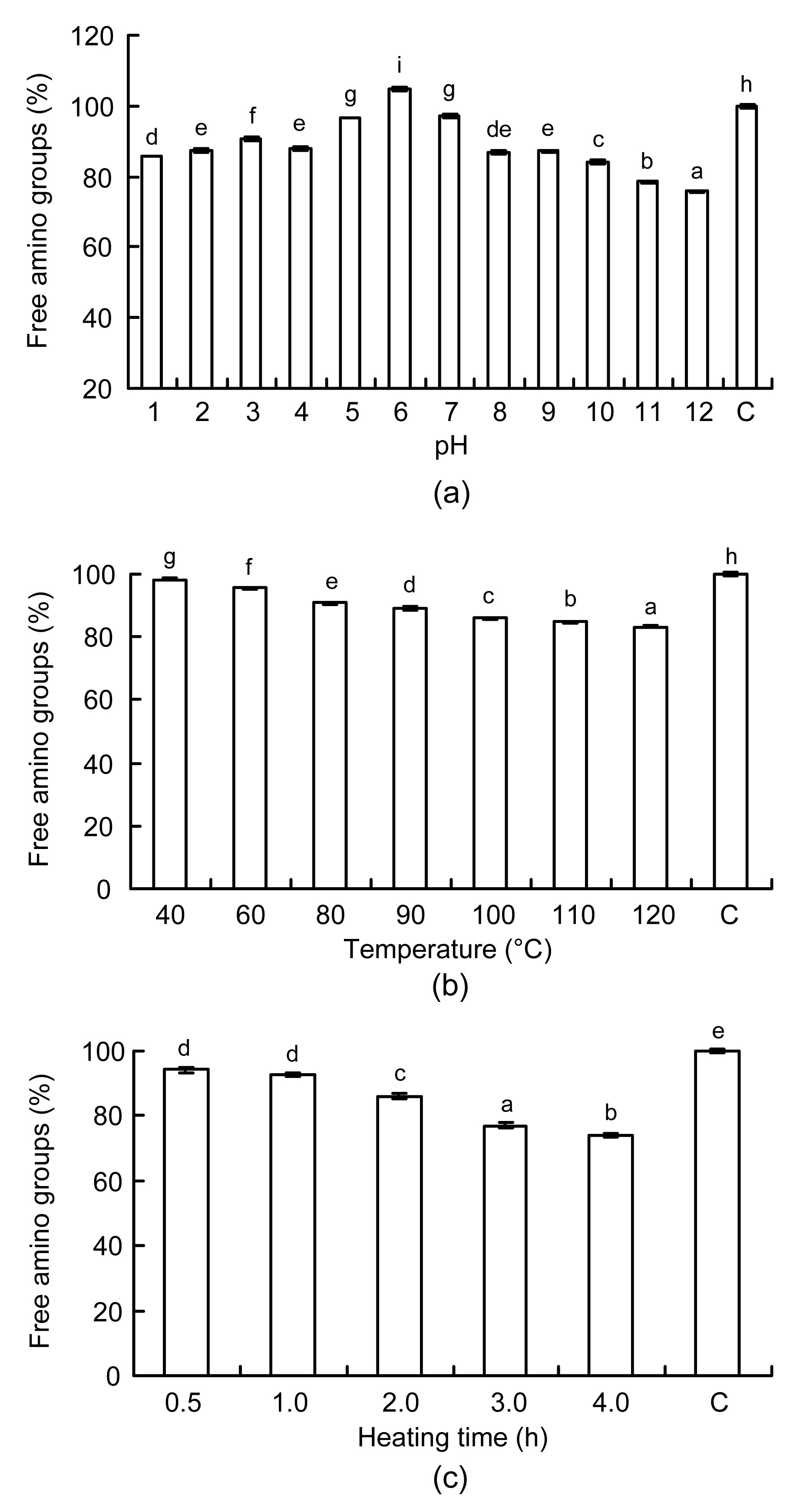
3.2. Effects of heat treatment on the free amino groups of bovine casein-derived ACE inhibitory peptides
To assay the availability of free amino groups, the OPA method with some modifications was used to measure the content of free amino groups in bovine casein-derived ACE inhibitory peptides. Changes in free amino group content of bovine casein-derived peptides at different pH levels (1.0 to 12.0) at 100 °C for 1 h are shown in Fig.
2a. In the pH range of 5.0 to 7.0, the content was maintained at 96.60%–104.69% of that of unheated samples. However, with increasing acidic and alkaline conditions, the content of free amino groups gradually declined (
P<0.05). When heated at pH 1.0 and 12.0, bovine casein-derived peptides lost 14.28% and 24.19% of free amino groups, respectively. This indicated that extremely acidic or alkaline heat treatment could destroy some amino groups of ACE inhibitory peptides. This was probably caused by chemically oxidative deamination of amino groups of the peptides during the heat treatment combined with acidic or alkaline conditions. Furthermore, alkaline heat treatment consumed more amino groups of the bovine casein-derived peptides than acidic heat treatment. Alkaline heating may have accelerated the condensation of amino groups with carbonyl groups of small reducing sugars present in the peptide samples.
Fig.2
Changes in free amino groups of ACE inhibitory peptides
(a) At different pH levels (1.0–12.0), 100 °C for 1 h of heat treatment; (b) At different temperatures (40–120 °C), pH 7.0 for 2 h; (c) For different heating time (0–4 h) at 100 °C and pH 7.0. C denotes the ACE inhibitory peptides without heat treatment. Values are expressed as mean±SD, n=3. Different letters above the bars are significantly different (P<0.05)
Changes in the free amino group content in the bovine casein-derived peptides at different temperatures (40 to 120 °C) at pH 7.0 for 2 h are shown in Fig.
2b. The free amino group content ranged from 98.27% to 83.29% with increasing temperature (
P<0.05). This suggests that high temperature treatment might also promote oxidative deamination of amino groups, leading to the loss of free amino groups from bovine casein-derived peptides.
Changes in the free amino group content of ACE inhibitory peptides derived from bovine casein heated for different time periods (0 to 4 h) at 100 °C and pH 7.0 are shown in Fig.
2c. After half an hour, the content decreased by 5.90%, compared to that of unheated peptides. There were no significant differences in the free amino group content (
P>0.05) within the first 1 h. The loss of free amino groups increased when the heating time was prolonged to 4 h (
P<0.05). Rada-Mendoza et al. (
2006) found that considerable losses in free amino groups could be detected in β-lactoglobulin heated for 120 h at 80 °C. These results indicate that prolonged heating time under high temperature conditions may increase or decrease the free amino group content of ACE inhibitory peptides of bovine casein.
3.3. Effects of heat treatment on the color of bovine casein-derived ACE inhibitory peptides
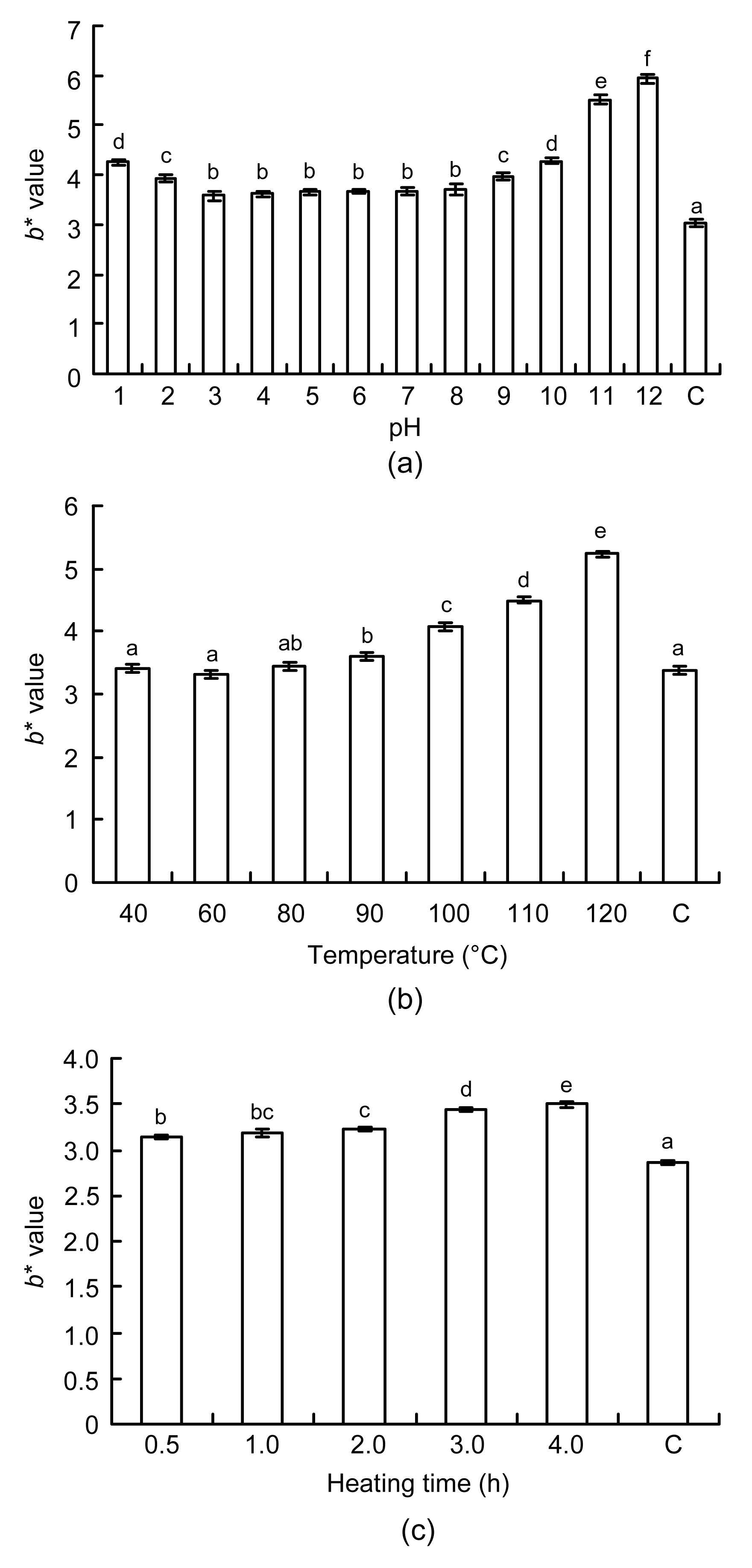
Color can be a visual expression of the extent of a reaction. When casein-derived ACE inhibitory peptides are dissolved in distilled water, the color of the solution is light yellow. However, heat treatment conditions probably promote the peptides to react with small amounts of sugars to form browning products. This would potentially darken the color of the peptide solution. Therefore, by detecting color changes, it may be possible to assess how different heat treatments affect ACE inhibitory peptides.
The changes in color of ACE inhibitory peptides under different pH values (1.0 to 12.0) at 100 °C for 1 h are shown in Fig.
3a. The
b
* values of ACE inhibitory peptides across this pH range were significantly different (
P<0.05) to that of unheated peptides. This showed that heat treatment could darken the color of ACE inhibitory peptide solutions. The
b
* values of peptides heated at pH levels of 1.0 and 2.0 were higher than those of peptides heated in the pH range of 3.0 to 8.0 (
P<0.05). This was probably due to strong acidity contributing to the formation of more chromatic compounds by chemical oxidation and aggregation of ACE inhibitory peptides. However, when the peptides were heated in pH range of 3.0 to 8.0, there were no significant differences in the
b
* values (
P>0.05). Nevertheless, ACE inhibitory peptides became darker and browner with increasingly alkaline pH values (9.0 to 12.0). Under alkaline heat treatment, amino groups of ACE peptides may have easily reacted with a few carbonyl groups present in the peptide samples to form browning products of the Maillard reaction, such as melanoidin. Thus, excessive alkaline heat treatment markedly changed the color of the bovine casein ACE inhibitory peptides.
Fig.3
Changes in the color of ACE inhibitory peptides
(a) At different pH levels (1.0–12.0), 100 °C for 1 h of heat treatment; (b) At different temperatures (40–120 °C), pH 7.0 for 2 h; (c) For different heating times (0–4 h) at 100 °C and pH 7.0. C denotes the ACE inhibitory peptides without heat treatment. Values are expressed as mean±SD, n=3. Different letters above the bars are significantly different (P<0.05)
Compared to unheated peptides, the
b
* values of ACE inhibitory peptides did not differ significantly in the temperature range of 40 to 80 °C (
P>0.05) (Fig.
3b). However, there was a significant increase in the
b
* value of samples heated from 90 to 120 °C (
P<0.05). Changes in the color of ACE inhibitory peptides under different heating times (0.5–4.0 h) at 100 °C and pH 7.0 are shown in Fig.
3c. The color of the peptides derived from bovine casein gradually darkened as the heating time increased up to 4 h (
P<0.05). These results proved that high temperature and prolonged heating time can accelerate the browning intensity due to the Maillard reaction in ACE inhibitory peptides.
In this study, the loss of ACE inhibitory activity of bovine casein-derived peptides was positively correlated with increasing dark color during heat treatment. Li et al. (
2010) reported that the ACE inhibitory activities of extracts from Chinese commercial fermented soy paste were significantly correlated with their color. Okamoto et al. (
1995) showed that soy sauces with light color showed lower ACE inhibitory activity than those with dark color. Thus, the ACE inhibitory activities of food ingredients may be correlated with their color.
3.4. Cell viability of bovine casein-derived ACE inhibitory peptides
Although some bioactive peptides have great potential as food functional ingredients, it is important to consider their safety and toxicity to ensure that they are suitable for being used in functional foods. To evaluate the cytotoxic effects of bovine casein-derived ACE inhibitory peptides, MTT assay was used to determine the cell viability. The effects of bovine casein-derived ACE inhibitory peptides unheated or heated for 5 h on the cell viability of Caco-2 and ECV-304 cell lines are shown in Fig.
4.
Fig.4
Effects of ACE inhibitory peptides unheated or heated at 100 °C and pH 7.0 for 5 h on cell viability
(a) Caco-2 cell line; (b) ECV-304 cell line. Values are expressed as mean±SD, n=6. Different letters above the bars are significantly different (P<0.05)
Our results demonstrated that there were no cytotoxic effects of bovine casein-derived ACE inhibitory peptides on the two types of human cell lines (Caco-2 and ECV-304) after incubation for 48 h in the DMEM solution containing a range of concentrations of the peptides from 0.01 to 0.20 mg/ml. Similar results were obtained by Qian et al. (
2007), who proved that ACE inhibitory peptides from bullfrog (
Rana catesbeiana Shaw) muscle protein showed no cytotoxicity to human embryonic lung fibroblast cell line MRC-5. There were also no deleterious cytochemical consequences (apoptotic, anti-proliferative or general cytotoxic effects) when a range of casein phosphopeptide preparations were added to human cancer cell lines (HL-60 and Caco-2 cells) and non-malignant polymorphonuclear leukocytes (Hartmann and Meisel,
2004).
When Caco-2 cells were incubated in DMEM with unheated ACE inhibitory peptide samples at concentrations from 0.01 to 0.20 mg/ml, no significant differences in cytotoxicity were found (
P>0.05) (Fig.
4a). Heating of ACE inhibitory peptide samples for 5 h increased the viability of Caco-2 cells (
P<0.05) compared to unheated samples. The viability of Caco-2 cells was significantly increased when the concentration of peptides heated for 5 h ranged from 0.05 to 2.0 mg/ml (
P<0.05). The results indicate that heated ACE inhibitory peptides were probably decomposed into some smaller fractions which could be absorbed by the cells and which stimulated cell proliferation by enhancing DNA synthesis (Azuma et al.,
1989; MacDonald et al.,
1994).
However, there were no significant differences in the viability of ECV-304 cells between unheated and heated ACE inhibitory peptide samples (
P>0.05) across a range of concentrations from 0.01 to 0.20 mg/ml (Fig.
4b). This result showed that heated ACE inhibitory peptides promoted proliferation of Caco-2 cells, but not of ECV-304 cells. This may be due to the different cell lines having different sensibility to the anti-proliferative or stimulating effects of the ACE inhibitory peptides (Phelan et al.,
2009).
4. Conclusions
Bovine casein-derived ACE inhibitory peptides were studied for the stability of their ACE inhibitory activity. Their free amino group content, color, and effects on cell viabilities of Caco-2 and ECV-304 were measured after heat treatment. Bovine casein-derived peptides could maintain ACE inhibitory activity with thermal treatment under acid, neutral, and weak alkaline conditions (pH 1.0–8.0), while strong alkaline conditions (pH 9.0–12.0) led to the loss of ACE inhibitory activity of the peptides. High temperatures (110–120 °C) also resulted in the loss of ACE inhibitory activity. The free amino group content of ACE inhibitory peptides decreased markedly under thermal treatment in acidic or alkaline conditions. Alkaline heat treatment (pH 9.0–12.0) greatly darkened the color of the peptides. Both high temperature and prolonged heating time led to the loss of free amino groups and dark color of ACE inhibitory peptides. However, heated ACE inhibitory peptides showed no cytotoxicity to Caco-2 or ECV-304 cells. Therefore, when ACE inhibitory peptides are applied as functional food ingredients, it is necessary to avoid severe heat treatments during food processing and storage.
* Project supported by the Innovative Research Team of Higher Education of Heilongjiang Province (No. 2010td11), the National Natural Science Foundation of China (No. 31000801), the National Key Technology R&D Program of China during the 12th Five-Year Plan Period (No. 2013BAD18B06), and the 2009 Doctoral Science Research of Northeast Agricultural University, ChinaCompliance with ethics guidelines Wei WU, Pan-pan YU, Feng-yang ZHANG, Hong-xia CHE, and Zhan-mei JIANG declare that they have no conflict of interest.References
[1] Akıllıoğlu, H.G., Karakaya, S., 2009. Effects of heat treatment and
in vitro digestion on the angiotensin converting enzyme inhibitory activity of some legume species.
Eur Food Res Technol, 229(6):915-921.

[2] Atkinson, A.B., Morton, J.J., Brown, J.J., 1980. Captopril in clinical hypertension. Changes in components of renin-angiotensin system and in body composition in relation to fall in blood pressure with a note on measurement of angiotensin II during converting enzyme inhibition.
Br Heart J, 44(3):290-296.

[3] Azuma, N., Nagaune, S., Ishino, Y., 1989. DNA-synthesis stimulating peptides from human β-casein.
Agric Biol Chem, 53(10):2631-2634.
[4] Boye, J.I., Ma, C.Y., Ismail, A., 1997. Molecular and microstructural studies of thermal denaturation and gelation of β-lactoglobulins A and B.
J Agric Food Chem, 45(5):1608-1618.
[5] Chen, G.W., Tsai, J.S., Pan, B.S., 2007. Purification of angiotensin I-converting enzyme inhibitory peptides and antihypertensive effect of milk produced by protease-facilitated lactic fermentation.
Int Dairy J, 17(6):641-647.
[6] Chen, J.W., Wang, Y.M., Zhong, Q.X., 2012. Purification and characterization of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide derived from enzymatic hydrolysate of grass carp protein.
Peptides, 33(1):52-58.
[7] Church, F.C., Swaisgood, H.E., Porter, D.H., 1983. Spectrophotometric assay using
o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins.
J Dairy Sci, 66(6):1219-1227.
[8] Costa, E.L., Gontijo, J.A.D., Netto, F.M., 2007. Effect of heat and enzymatic treatment on the antihypertensive activity of whey protein hydrolysates.
Int Dairy J, 17(6):632-640.
[9] Cushman, D.W., Cheung, H.S., 1971. Spectrophotometric assay and properties of the angiotensin converting enzyme of rabbit lung.
Biochem Pharmacol, 20(7):1637-1648.
[10] Ferreira, S.H., 1965. A bradykinin-potentiating factor (BPF) present in the venom of
Bothrops jararaca
.
Br J Pharmacol Chemother, 24(1):163-169.
[11] Hartmann, R., Meisel, H., 2004. Caseinophosphopeptides and their cell modulting potential.
Biofactors, 21(1-4):73-78.
[12] Hwang, J.S., 2010. Impact of processing on stability of angiotensin I-converting enzyme (ACE) inhibitory peptides obtained from tuna cooking juice.
Food Res Int, 43(3):902-906.
[13] Ichimura, T., Hu, J.N., Aita, D.Q., 2003. Angiotensin I-converting enzyme inhibitory activity and insulin secretion stimulative activity of fermented fish sauce.
J Biosci Bioeng, 96(5):496-499.
[14] Jang, A., Lee, M., 2005. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates.
Meat Sci, 69(4):653-661.
[15] Jang, A., Jo, C., Lee, M., 2007. Storage stability of the synthetic angiotensin converting enzyme (ACE) inhibitory peptides separated from beef sareoplasmic protein extracts at different pH, temperature, and gastric digestion.
Food Sci Biotechnol, 16(4):572-575.
[16] Jiang, Z.M., Tian, B., Brodkorb, A., 2010. Production, analysis and
in vivo evaluation of novel angiotensin-I-converting enzyme inhibitory peptides from bovine casein.
Food Chem, 123(3):779-786.
[17] Korhonen, H., Pihlanto-Leppala, A., Rantamaki, P., 1998. Impact of processing on bioactive proteins and peptides.
Trends Food Sci Technol, 9(8-9):307-319.
[18] Lee, J.K., Hong, S., Jeon, J.K., 2009. Purification and characterization of angiotensin I converting enzyme inhibitory peptides from the rotifer,
Brachionus rotundiformis
.
Bioresource Technol, 100(21):5255-5259.
[19] Li, F.J., Yin, L.J., Cheng, Y.Q., 2010. Angiotensin I-converting enzyme inhibitory activities of extracts from commercial Chinese style fermented soypaste.
Jpn Agric Res Q, 44(2):167-172.
[20] Li, G.H., Le, G.W., Shi, Y.H., 2004. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects.
Nutr Res, 24(7):469-486.
[21] Lpez-Fandino, R., Otte, J., van Camp, J., 2006. Physiological, chemical and technological aspects of milk-protein-derived peptides with antihypertensive and ACE-inhibitory activity.
Int Dairy J, 16(11):1277-1293.
[22] MacDonald, R., Thornton, W.H., Marshall, R.A., 1994. A cell culture model to identify biologically active peptides generated by bacterial hydrolysis of casein.
J Dairy Sci, 77(5):1167-1175.
[23] Okamoto, A., Hanagata, H., Matsumoto, E., 1995. Angiotensin I converting enzyme inhibitory activities of various fermented foods.
Biosci Biotechnol Biochem, 59(6):1147-1149.
[24] Ondetti, M.A., Cushman, D.W., 1982. Enzyme of the rennin-angiotensin system and their inhibitors.
Annu Rev Biochem, 51(1):283-308.
[25] Oshima, G., Shimabukuro, H., Nagasawa, K., 1979. Peptide inhibitors of angiotensin I-converting enzyme in digests of gelatin by bacterial collagenase.
Biochim Biophys Acta Enzymol, 566(1):128-137.
[26] Phelan, M., Aherne, A., FitzGerald, R.J., 2009. Casein-derived bioactive peptides: biological effects, industrial uses, safety aspects and regulatory status.
Int Dairy J, 19(11):643-654.
[27] Qian, Z.J., Jung, W.K., Lee, S.H., 2007. Antihypertensive effect of an angiotensin I-converting enzyme inhibitory peptide from bullfrog (
Rana catesbeiana Shaw) muscle protein in spontaneously hypertensive rats.
Process Biochem, 42(10):1443-1448.
[28] Rada-Mendoza, M., Villamie, M., Molina, E., 2006. Effects of heat treatment and high pressure on the subsequent lactosylation of beta-lactoglobulin.
Food Chem, 99(4):651-655.
[29] Rokar, R., Simončič, Z., Gartner, A., 2009. Stability of new potential ACE inhibitor in the aqueous solutions of different pH.
J Pharm Biomed Anal, 49(2):295-303.
[30] Sheih, I.C., Fang, T.J., Wu, T.K., 2009. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste.
Food Chem, 115(1):279-284.
[31] Tovar-Prez, E.G., Guerrero-Legarreta, I., Farres-Gonzalez, A., 2009. Angiotensin I-converting enzyme-inhibitory peptide fractions from albumin 1 and globulin as obtained of amaranth grain.
Food Chem, 116(2):437-444.
[32] Wu, J.P., Ding, X.L., 2002. Characterization of inhibition and stability of soy-protein-derived angiotensin I-converting enzyme inhibitory peptides.
Food Res Int, 35(4):367-375.
[33] Zhang, J.H., Tatsumi, E., Ding, C.H., 2006. Angiotensin I-converting enzyme inhibitory peptides in douchi, a Chinese traditional fermented soybean product.
Food Chem, 98(3):551-557.



Open peer comments: Debate/Discuss/Question/Opinion
<1>