1. Introduction
The uterus is a major female hormone-responsive organ necessary for many pivotal reproductive events, such as embryo implantation, decidualization, placentation, and labor (Dey,
2004; Fernandez-Valdivia et al.,
2007; Franco et al.,
2008; Abdallah et al.,
2012; Kong et al.,
2012). Endometrial tissues undergo remarkable periodic growth, degeneration, and leucocytosis of the epithelial cells during the estrous cycle. These histological events are controlled by ovarian steroids. In addition, many of these processes are considered to be regulated by cytokines and growth factors, several of which are members of the transforming growth factor-β (TGF-β) superfamily (Liu et al.,
2004; Omwandho et al.,
2010; Bukowska et al.,
2011; Tanwar and McFarlane,
2011; Argañaraz et al.,
2013).
The bone morphogenetic protein (BMP) family is the largest subgroup of ligands in the TGF-β superfamily, which is closely associated with biological processes and cellular events involved in uterine cell proliferation, differentiation, apoptosis, and tissue remodeling (Shimasaki et al.,
2004; Shimizu et al.,
2004; Jones et al.,
2006). More than 20 BMP isoforms have been identified, and they can be divided into four distinct subgroups based on the properties of their binding to type I receptors, including BMP2/4, BMP5/6/7/8, growth/differentiation factor (GDF)-5/6/7, and BMP9/10. Homo- or hetero-dimers of the BMP family ligands bind to and phosphorylate two types of serine/threonine kinase receptors to form complexes of transmembrane type I and type II receptors, which subsequently transmit signals from the cell surface to the cytoplasm (Aoki et al.,
2001; Hu et al.,
2003; Cai et al.,
2012).
The Smad-dependent pathway involves canonical BMP signaling and mediates signals from the cytoplasm to the nucleus (Wen et al.,
2011). SMAD proteins, the major intracellular signal transducers for the BMP family receptors, can be classified into three types: receptor-regulated SMADs (R-SMADs: SMAD1, SMAD5, and SMAD8), inhibitory SMADs (I-SMADs: SMAD6 and SMAD7), and a common-mediator SMAD (SMAD4) (Massague et al.,
2005; Miyazono et al.,
2010; Cai et al.,
2012; Pangas,
2012). Once a ligand-receptor complex is formed, type-I receptors phosphorylate down-stream R-SMAD proteins. Phosphorylated R-SMADs form heteromeric complexes with SMAD4 (Co-SMAD) and translocate to the nucleus to regulate BMP-stimulated alterations in target gene expression through interaction with other factors, including transcription factors and transcriptional coactivators or corepressors (Xia et al.,
2005; Bruce and Sapkota,
2012).
Several molecular and functional studies have suggested the presence of BMP family members in the mammalian uterus (Paria et al.,
2001; Erickson et al.,
2004; Lee et al.,
2007).
In situ hybridization results revealed that
Bmp7 was highly expressed in the endometrium of the non-pregnant rat uterus (Erickson et al.,
2004). A low level of
Bmp5 mRNA has been detected in the stroma close to the myometrium and in the myometrial connective tissue (Paria et al.,
2001).
Bmpr mRNA appeared to show dynamic expression (Erickson et al.,
2004), whereas
Smad4 mRNA was constitutively expressed in the glandular and luminal epithelium throughout the estrous cycle (Liu et al.,
2004). The differential distribution and expression of BMP family members in the uterus indicate their differing roles in the regulation of uterine function and development. There is an increasing evidence that BMP2 affects proliferation, differentiation, and apoptosis of endometrial cells (Lee et al.,
2007; Franco et al.,
2011) and that BMP4 may play an important role in the maintenance of the uterine endometrium (Tanwar and McFarlane,
2011). Although a series of studies have focused on BMP signaling, there is a lack of quantitative descriptions of BMP family members in the mouse uterus. To gain more experimental clues clarifying the molecular mechanisms of BMP signaling during the estrous cycle, we analyzed the temporal and spatial expression patterns of BMP family members by real-time polymerase chain reaction (PCR) and immunohistochemistry.
2. Materials and methods
2.1. Experimental animals and sample preparation
A total of 30 female Swiss ICR mice of similar age and weight were purchased from the Laboratory Animal Center of Nanjing Medical University, China. They were raised in an animal room with controlled temperature (20–25 °C) and lighting (12-h light/dark cycle), and were provided with commercial feed and tap water
ad libitum. The various stages of the estrous cycle were monitored by vaginal smears. The body weights of the mice were recorded immediately before they were sacrificed. Uterine samples (at least seven mice per time point) were collected and dissected free of surrounding tissue. One uterine horn was snap frozen in liquid nitrogen immediately after collection and stored at −80 °C for RNA extraction, and the other was fixed in 40 mg/ml paraformaldehyde at room temperature for immunolocalization of BMP2 protein. All experiments were carried out in accordance with the principles for the Care and Use of Laboratory Animals approved by the Institutional Animal Care and Use Committee of Nanjing Agricultural University, China.
2.2. RNA isolation and reverse transcription
Total RNA was extracted from all samples using a commercial RNA isolation kit (TaKaRa code: D9108A, Dalian, China) according to the manufacturer’s instructions. Total RNA was dissolved in 60 μl RNase-free deionized water (dH
2O) and stored at −80 °C. Subsequently, 1 μg of total RNA of high integrity (not degraded) and high purity (no contaminants) was reverse-transcribed into complementary DNA (cDNA) using reverse transcription reagent kits (TaKaRa code: RR047A, Dalian, China). Each PCR reaction contained 2.0 μl 5× genomic DNA (gDNA) eraser buffer, 1.0 μl gDNA eraser, 1 μg total RNA, 1.0 μl PrimeScript™ RT enzyme mix 1, 1.0 μl RT primer mix, 1.0 μl RT primer mix, 4.0 μl 5× PrimeScript™ buffer 2, and sufficient RNase-free dH
2O in a total volume of 20 μl. Reverse transcription cycle conditions were: 42 °C for 2 min and 37 °C for 15 min. The reaction was terminated by incubation at 85 °C for 5 s and cooling at 4 °C. The cDNA was stored at −80 °C or used for real-time PCR immediately.
2.3. Real-time PCR
The mRNA expression patterns of BMP2, BMP4, and components of the BMP signaling pathway were assessed using a commercial real-time PCR kit (TaKaRa code: RR820A, Dalian, China) on an ABI StepOne™ real-time PCR machine (Applied Biosystems, Life Technologies Corporation, Grand Island, NY, USA). The primer pairs used for amplification of the target genes are listed in Table
1. The PCR amplification was performed in a total volume of 20 μl containing 10 μl SYBR Premix Ex Taq II, 6.0 μl RNase-free dH
2O, 2 μl cDNA, 0.4 μl ROX reference dye, and 0.8 μl each of forward and reverse primer pairs. Each PCR cycle included a denaturation step at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and extension at 60 °C for 34 s, and a dissociation step consisting of 95 °C for 15 s, 60 °C for 1 min, and 95 °C for 15 s. The real-time PCR was performed with three technical replicates per animal sample. Finally, the relative expression of each gene was standardized to glyceraldehyde-3-phosphate dehydrogenase (
Gapdh).
Table 1
Primer sequences, GenBank accession Nos., and product sizes of genes selected for real-time PCR
| Target gene |
Accession No. |
Primer sequence (5'→3') |
Product size (bp) |
|
Gapdh
|
NM_008084.2 |
F: CATGTTCCAGTATGACTCCACTC |
136 |
| R: GGCCTCACCCCATTTGATGT |
|
Bmp2
|
NM_007553.2 |
F: TGACTGGATCGTGGCACCTC |
112 |
| R: CAGAGTCTGCACTATGGCATGGTTA |
|
Bmp4
|
NM_007554.2 |
F: TTTGTTCAAGATTGGCTCCCAAG |
101 |
| R: AAACGACCATCAGCATTCGGTTA |
|
Bmpr1a
|
NM_009758.4 |
F: ATGGCCATTGCTTTGCCATTA |
126 |
| R: TCCTGCGTAGCTGGGCTTTC |
|
Bmpr1b
|
NM_007560.3 |
F: CTACGTTGTAAATGCCACCACCA |
118 |
| R: AGGTGACAACAGGCATTCCAGA |
|
Bmpr2
|
NM_007561.3 |
F: CAAATATCCTGGATGGCAGCAGTA |
141 |
| R: TGGAGATGACCCAGGTGGAC |
|
Smad1
|
NM_008539.3 |
F: ACCTGTGGCTTCCGTCTC |
97 |
| R: ATCGTGGCTCCTTCGTC |
|
Smad4
|
NM_008540.2 |
F: CAGCACTACCACCTGGACTGGA |
145 |
| R: CTGGAATGCAAGCTCATTGTGAA |
|
Smad5
|
NM_001164041.1 |
F: AAGTAGATTCTGCCTGGGATT |
197 |
| R: AGACGGTGGTGGGATGG |
|
Smad8
|
NM_019483.5 |
F: ATCCCTGGCAATCTGTA |
169 |
| R: CCCTGGCTGTCCTGTAA |
2.4. Immunolocalization of BMP2 protein
Immunohistochemistry was carried out using the streptavidin-biotin peroxidase complex (SABC) method to examine the immunolocalization of BMP2 protein in the mouse uterus during the estrous cycle. Paraffin-embedded uteri were cut into 6-μm-thick sections and mounted on slides. Sections were deparaffinised and hydrated by a consecutive series of xylene and ethanol, and treated with 9 mg/ml hydrogen peroxide to inactivate endogenous peroxidase activity. Sections were then incubated with 50 mg/ml bovine serum albumin (BSA) over 1 h to block non-specific staining. Subsequently, sections were incubated with a rabbit polyclonal antibody to BMP2 (Abcam Inc., Cambridge, MA, USA) at a dilution of 1:400 overnight at 4 °C, and then incubated with a secondary antibody, goat anti-rabbit IgG, diluted in phosphate buffer solution (PBS) over 2 h in a wet box at room temperature. Finally, immunoreactive signals were visualized by addition of diaminobenzidine substrate (DAB; Sigma Chemical Co., USA). Negative control sections were incubated with normal rabbit serum omitting the primary antibody. To distinguish the staining intensity of BMP2 protein among the different uterine structural components and cell types, sections were examined using a method similar to that described previously (Shi et al.,
2004; Ding et al.,
2012; Li et al.,
2014): −, no staining; +, weak staining; ++, moderate staining; +++, strong staining. Relative levels of immunostaining were evaluated at least three times for each sampling stage.
2.5. Statistical analysis
All data were collected using Origin Pro 8.0 software (Origin Lab Corporation, USA). Data are presented as mean±standard error of the mean (SEM). For statistical evaluation of data, one-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test was applied to detect differences among stages for each gene. Paired-sample
t-tests were performed for two-gene comparisons. Means identified with the same letter are not significantly different (
P>0.05).
3. Results
3.1. Body weights of mice during the estrous cycle
As shown in Table
2, the body weight of each mouse was nearly 30 g, and there were no significant differences among different stages of the estrous cycle.
Table 2
Body weights of mice during the estrous cycle
| Stage of estrous cycle |
Number of animals |
Body weight* (g) |
| P |
7 |
28.47±0.59 |
| E |
7 |
30.46±0.62 |
| M |
8 |
28.84±0.93 |
| D |
8 |
29.72±0.81 |
*Data are expressed as mean±SEM
P: proestrus; E: estrus; M: metestrus; D: diestrus
3.2. mRNA expression patterns of Bmp2, Bmp4, and components of the signaling pathway during the estrous cycle
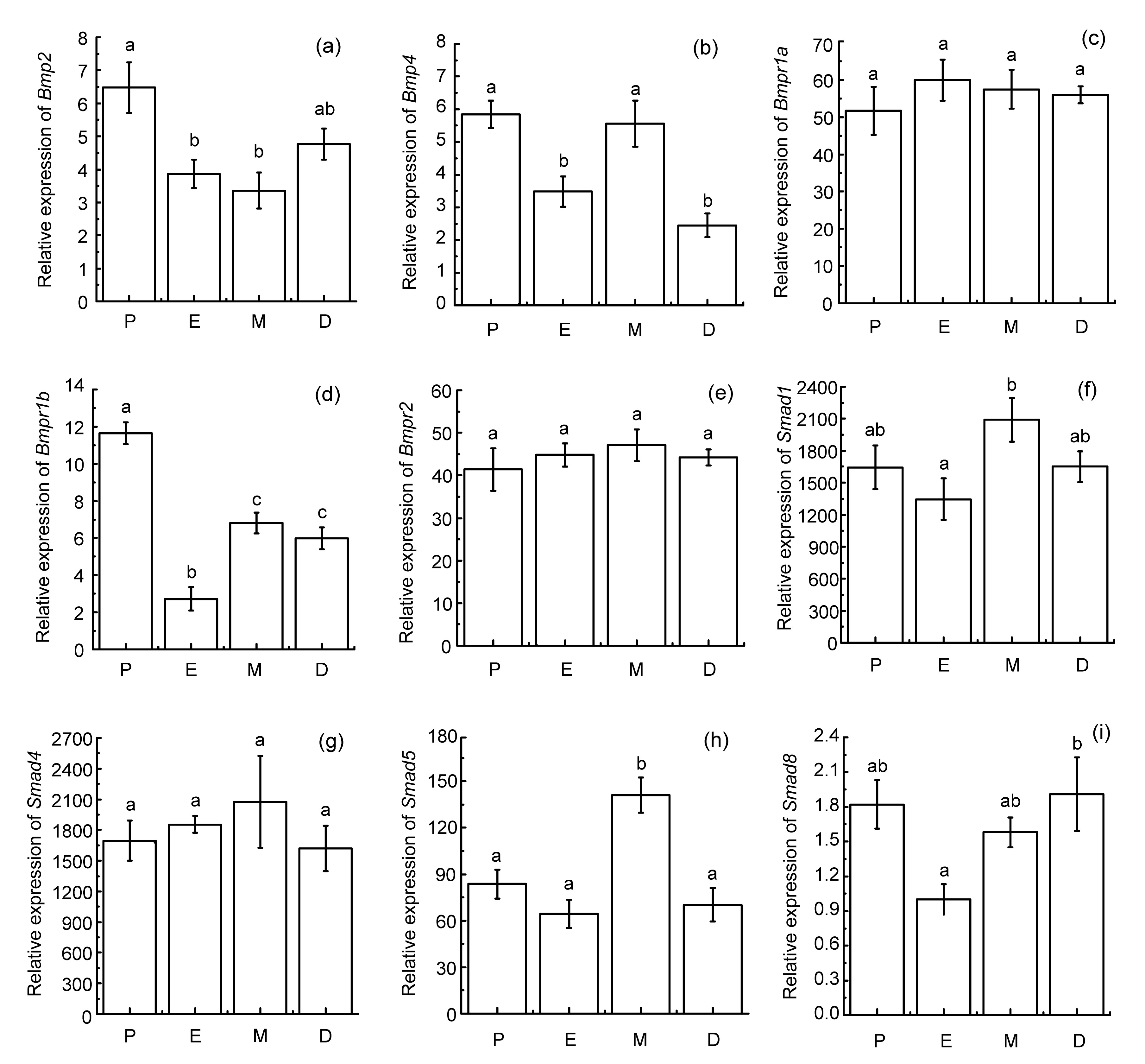
To investigate the mRNA levels of BMP family members, real-time PCRs were performed in the mouse uterus during the estrous cycle. Our results showed that different members of the BMP family were differentially expressed in all studied stages of the estrous cycle.
The expression level of
Bmp2 was significantly higher at proestrus (P) than at estrus (E) and metestrus (M) (
P<0.05; Fig.
1). The relative abundance of
Bmp4 showed significant fluctuations, strongly decreasing at estrus and diestrus (D) (
P<0.05), and strongly increasing at metestrus (
P<0.05). Paired-sample
t-tests revealed that there were no statistically significant differences between the expression levels of
Bmp2 and
Bmp4.
Fig.1
Changes in mRNA levels for Bmp2, Bmp4, and components of the signaling pathway throughout the estrous cycle
Uterine samples were collected at various stages of the estrous cycle (P: proestrus; E: estrus; M: metestrus; D: diestrus). Total RNA was extracted from all samples, and 1 μg total RNA was analyzed by real-time PCR. The relative expression of each gene was standardized to glyceraldehyde-3-phosphate dehydrogenase (Gapdh). Then, all data were normalized on the basis of level of Smad8 mRNA at estrus. All results were presented as mean±SEM (at least seven mice each stage). Means identified with the same letter are not significantly different (P>0.05)
The expression levels of
Bmpr1a and
Bmpr2 remained unchanged during an entire estrous cycle. However, the level of
Bmpr1b mRNA decreased significantly at estrus (
P<0.05), increasing subsequently at metestrus (
P<0.05). Furthermore, the level of
Bmpr1b mRNA was significantly lower compared with that of
Bmpr1a and
Bmpr2 mRNA at the corresponding stages (
P<0.05).
The expression level of
Smad1 did not change significantly at proestrus or estrus, whereas it increased markedly at metestrus (
P<0.05). The relative quantity of
Smad4 did not change substantially throughout the estrous cycle. The expression level of
Smad5 was significantly higher at metestrus than at other stages (
P<0.05). The relative quantity of
Smad8 was lower at estrus than at diestrus (
P<0.05). In addition, the expression levels of
Smad1 and
Smad5 were significantly higher than that of
Smad8 (
P<0.05).
3.3. Immunolocalization of BMP2 protein during the estrous cycle
To provide more evidence illustrating the important role of BMP signaling during the estrous cycle, the BMP2 protein in the mice uterus was examined using immunohistochemistry. Our results showed that BMP2 protein was detected throughout the estrous cycle (Fig.
2). Specifically, it was moderately immunolocalized to the uterine luminal and glandular epithelial cells at proestrus (Figs.
2a and 2e; Table
3), estrus (Figs.
2b and 2f; Table
3), and diestrus (Figs.
2d and 2h; Table
3), whereas it was weak at metestrus (Figs.
2c and 2g; Table
3). Furthermore, individual stromal cells and myometrium were weakly stained BMP2 protein positive (Fig.
2; Table
3). No specific staining was detected in the negative control sections (Figs.
2i and 2j).
Fig.2
BMP2 protein in the mouse during the estrous cycle: low magnification (a–d) and high magnification (e–h)
Uterine sections were immunostained with a rabbit polyclonal antibody to BMP2 and counterstained with haematoxylin. A positive reaction was observed as brown staining and the counterstaining background appeared blue. BMP2 protein was moderately immunolocalized to the uterine luminal and glandular epithelial cells at proestrus (a, e), estrus (b, f), and diestrus (d, h), whereas it was weak at metestrus (c, g). Furthermore, individual stromal cells and myometrium were weakly stained BMP2 protein positive. Negative control sections showed no specific staining (i, j). LEC: luminal epithelial cells; GEC: glandular epithelial cells; SC: stromal cells; ICM: inner circular smooth muscle layer; OLM: outer longitudinal smooth muscle layer. Scale bar: 10 μm (Note: for interpretation of the references to color in this figure legend, the reader is referred to the web version of this article)
Table 3
Relative abundances of BMP2 protein in the mouse uterus during the estrous cycle
| Uterine structural component and cell type |
Staining intensity
|
| Proestrus |
Estrus |
Metestrus |
Diestrus |
| Glandular epithelial cell |
++ |
++ |
+ |
++ |
| Luminal epithelial cell |
++ |
++ |
+ |
++ |
| Myometrial cell |
+ |
+ |
+ |
+ |
| Stromal cell |
+ |
+ |
+ |
+ |
+: weak staining; ++: moderate staining; +++: strong staining
4. Discussion
Our study provided novel information regarding the expression of
Bmp2 and
Bmp4 members of the BMP family and components of the signaling pathway at the mRNA level in the mouse uterus during the estrous cycle. To minimize individual differences, female Swiss ICR mice of similar age and body weight were used. Our findings strongly suggest that the
Bmp2 and
Bmp4 signaling pathway might play key roles in the regulation of cyclic changes in the mouse uterus.
A wealth of evidence has indicated that the mRNAs encoding a number of BMP family members are abundantly and dynamically expressed in the murine uterus, and are associated with cell proliferation, differentiation, and apoptosis (Paria et al.,
2001; Erickson et al.,
2004; Liu et al.,
2004; Xia et al.,
2005; Li et al.,
2007; Large and DeMayo,
2012; Wetendorf and DeMayo,
2012; Nagashima et al.,
2013). In our study, we examined the mRNA levels of
Bmp2 and
Bmp4 during the estrous cycle. Our results showing that the expression level of
Bmp2 was significantly higher at proestrus than at estrus and metestrus were inconsistent with Erickson et al. (
2004). This inconsistency might be caused by differences between animals and experimental techniques used for study.
Bmp2 ablation affects proliferation, differentiation, and apoptosis in the decidualizing uterus (Lee et al.,
2007; Franco et al.,
2011). Results of
in vitro experiments had suggested that BMP2 promoted differentiation of endometrial stromal cells (Li et al.,
2007). Thus, the expression of
Bmp2 in the uterus implies that BMP2 might have regulatory functions in the degradation and remodeling of the endometrium during the estrous cycle. However, the specific mechanism underlying this effect needs confirmation. In comparison to
Bmp2, the relative abundance of
Bmp4 showed significant fluctuations, strongly decreasing at estrus and diestrus, and strongly increasing at metestrus. We speculate that the significant change in
Bmp4 expression might be tightly correlated with the cyclic changes in the endometrium. This suggestion is nearly identical to that proposed by Tanwar and McFarlane (
2011). Increasing evidence suggests that there is a subtle relationship between steroids and BMP signaling. Expression of uterine
Bmp7 was abolished in response to estradiol treatment in mice (Shimasaki et al.,
2004; Tanwar and McFarlane,
2011). Similarly, exogenous treatment with progesterone receptor antagonist RU486 has been shown to markedly down-regulate the level of
Bmp2 mRNA in the decidual uterus (Li et al.,
2007). Further studies are needed to confirm a functional link between steroids and BMP signaling in the uterus. Paired-sample
t-tests revealed that there were no statistically significant differences between the expression levels of
Bmp2 and
Bmp4, which suggests the possibility that both play important roles in the control of uterine functions. Collectively, these findings suggest that both
Bmp2 and
Bmp4 might contribute to the degradation and remodeling of the endometrium during the estrous cycle.
BMP receptors are essential mediators of BMP signal transduction from the cell surface to the cytoplasm. BMPR1A and BMPR1B are receptors specific for BMP2 and BMP4, whereas BMPR2 appears to bind exclusively to BMP ligands among all BMP receptors identified in mammals (Shimasaki et al.,
2004; Miyazono et al.,
2010). Although much is known about BMP receptors, little is known about their roles. In the current study, the mRNA levels of the three
Bmprs were confirmed by real-time PCR.
Bmpr1a and
Bmpr2 showed extremely stable expression during an entire estrous cycle. Our findings suggest that both
Bmpr1a and
Bmpr2 are constitutively expressed in the uterus. In contrast, we found that the level of
Bmpr1b mRNA appeared to change significantly throughout the estrous cycle. The stage-specific changes in
Bmpr1b mRNA expression might be closely associated with the growth and development of the endometrium (Kim et al.,
2003; Erickson et al.,
2004). In support of this deduction, mice deficient in
Bmpr1b exhibited a failure in endometrial gland formation and severe destruction of the uterine structure (Yi et al.,
2001). Furthermore, our results showed that the level of
Bmpr1b mRNA was significantly lower than that of
Bmpr1a and
Bmpr2 mRNA.
In situ hybridization results showed that
Bmpr1a and
Bmpr2 mRNA appeared to be widely presented in the endometrium and myometrium, whereas
Bmpr1b message was restricted to the luminal and glandular epithelial cells (Erickson et al.,
2004). Thus, epithelial restricted expression of
Bmpr1b could be one of the reasons for the low expression level of
Bmpr1b compared to
Bmpr1a and
Bmpr2. Our findings, together with the studies cited above, suggest that
Bmpr1b is essential for maintenance of normal uterine functions. Our results strongly suggest that three murine BMP receptors differentially contribute to the control of growth and development of the endometrium during the estrous cycle.
Smad1,
Smad5, and
Smad8 are
R-Smads mediating the intracellular signaling of BMP. Earlier studies have reported their critical functions in different models (Chen et al.,
1997; Chang et al.,
1999; Ying and Zhao,
2000; Huang et al.,
2009). However, there is a lack of understanding of the role of each
R-Smad in the mouse uterus during the estrous cycle. Our results showed that all the three
R-Smads were differentially expressed in the mouse uterus, demonstrating their differing roles during the estrous cycle. Further analysis revealed that the expression levels of
Smad1 and
Smad5 were significantly higher than that of
Smad8. Thus, we conjecture that
Smad1 and
Smad5 are the predominant
R-Smads involved in the regulation of uterine function during the estrous cycle. Our results revealed also that
Smad4 was highly expressed throughout the estrous cycle. Our results are largely similar to those of previous works (Lin et al.,
2004; Liu et al.,
2004). The marked expression of
Smad4 might be closely correlated with its molecular roles.
Smad4, a common partner for all
R-Smads, works through combination with
R-Smads to control gene transcription (Wan et al.,
2002; Shimasaki et al.,
2004). Combining the relative level of
Smad4 in the current study with previous results in rodents, we speculate that the persistent and high-level expression of
Smad4 in the uterus is an essential factor for mediation of BMP signal during the estrous cycle.
We investigated the immunolocalization of BMP2 protein using immunohistochemistry. Our observation that BMP2 protein was localized mainly in the uterine luminal and glandular epithelial cells throughout the estrous cycle appears to contrast with the results of Erickson et al. (
2004). This discrepancy might be a result of differences between animals and experimental techniques. Our findings demonstrated that both uterine luminal and glandular epithelium were primary targets for BMP signaling. Differences in the staining intensity of BMP2 protein were observed during different stages of the estrous cycle. Thus, we surmised that BMP2 protein contributed to the degradation and remodeling of the endometrium in a stage-specific manner.
5. Conclusions
In conclusion, we detected the temporal expression patterns of
Bmp2 and
Bmp4 members of the BMP family and components of the signaling pathway in the mouse uterus during the estrous cycle. Also, we examined the histological location of BMP2 protein using immunohistochemistry. Our results provide information about the variation in the mRNA levels of
Bmp2 and
Bmp4 and components of the BMP signaling pathway. Our findings also provide quantitative and useful information about the roles of endometrial BMP proposed and demonstrated by others, such as the degradation and remodeling of the endometrium.
Acknowledgements
We sincerely thank Dr. Reinhold J. HUTZ of the Department of Biological Sciences, University of Wisconsin-Milwaukee, USA, for reading the original manuscript and providing valuable suggestions.
* Project supported by the National Natural Science Foundation of China (No. 31172206) and the Grant-in-Aid for Innovative Training of Doctoral Students in Jiangsu Province (No. CXLX13-287), China Yan LI, Quan-wei WEI, Jian-gang FENG, Mu-lin XU, Rui-hua HUANG, and Fang-xiong SHI declare that they have no conflict of interest.References
[1] Abdallah, Y., Naji, O., Saso, S., 2012. Ultrasound assessment of the peri-implantation uterus: a review.
Ultrasound Obstet Gynecol, 39(6):612-619.


[2] Aoki, H., Fujii, M., Imamura, T., 2001. Synergistic effects of different bone morphogenetic protein type I receptors on alkaline phosphatase induction.
J Cell Sci, 114(8):1483-1489.
[3] Argañaraz, M.E., Apichela, S.A., Kenngott, R., 2013. Expression and localization of nodal in bovine oviduct and uterus during different functional stages of oestrus cycle and pregnancy.
Histochem Cell Biol, 139(1):89-97.
[4] Bruce, D.L., Sapkota, G.P., 2012. Phosphatases in SMAD regulation.
FEBS Lett, 586(14):1897-1905.
[5] Bukowska, D., Kempisty, B., Jackowska, M., 2011. Differential expression of epidermal growth factor and transforming growth factor beta isoforms in dog endometrium during different periods of the estrus cycle.
Pol J Vet Sci, 14(2):259-264.
[6] Cai, J., Pardali, E., Sanchez-Duffhues, G., 2012. BMP signaling in vascular diseases.
FEBS Lett, 586(14):1993-2002.
[7] Chang, H., Huylebroeck, D., Verschueren, K., 1999. Smad5 knockout mice die at mid-gestation due to multiple embryonic and extraembryonic defects.
Development, 126(8):1631-1642.
[8] Chen, Y., Bhushan, A., Vale, W., 1997. Smad8 mediates the signaling of the ALK-2 receptor serine kinase.
PNAS, 94(24):12938-12943.
[9] Dey, S.K., 2004. Focus on implantation.
Reproduction, 128(6):655-656.
[10] Ding, W., Zhang, W., Hui, F.M., 2012. Cell-specific expression and immunolocalization of nitric oxide synthase isoforms and soluble guanylyl cyclase α
1 and β
1 subunits in the ovary of fetal, neonatal and immature pigs.
Anim Reprod Sci, 131(3-4):172-180.
[11] Erickson, G.F., Fuqua, L., Shimasaki, S., 2004. Analysis of spatial and temporal expression patterns of bone morphogenetic protein family members in the rat uterus over the estrous cycle.
J Endocrinol, 182(2):203-217.
[12] Fernandez-Valdivia, R., Mukherjee, A., Amato, P., 2007. Progesterone-action in the murine uterus and mammary gland requires steroid receptor coactivator 2: Relevance to the human.
Front Biosci, 12(8-12):3640-3647.
[13] Franco, H.L., Jeong, J.W., Tsai, S.Y., 2008.
In vivo analysis of progesterone receptor action in the uterus during embryo implantation.
Semin Cell Dev Biol, 19(2):178-186.
[14] Franco, H.L., Dai, D., Lee, K.Y., 2011. Wnt4 is a key regulator of normal postnatal uterine development and progesterone signaling during embryo implantation and decidualization in the mouse.
FASEB J, 25(4):1176-1187.
[15] Hu, J., Zhang, Y.Q., Liu, X.P., 2003. Expression and localization of Smad1, Smad2 and Smad4 proteins in rat testis during postnatal development.
Asian J Androl, 5(1):51-55.
[16] Huang, Z., Wang, D., Ihida-Stansbury, K., 2009. Defective pulmonary vascular remodeling in
Smad8 mutant mice.
Hum Mol Genet, 18(15):2791-2801.
[17] Jones, R.L., Stoikos, C., Findlay, J.K., 2006. TGF-β superfamily expression and actions in the endometrium and placenta.
Reproduction, 132(2):217-232.
[18] Kim, J.G., Song, J.H., Vallet, J.L., 2003. Molecular characterization and expression of porcine bone morphogenetic protein receptor-IB in the uterus of cyclic and pregnant gilts.
Biol Reprod, 68(3):735-743.
[19] Kong, S., Zhang, S., Chen, Y., 2012. Determinants of uterine aging: lessons from rodent models.
Sci China Life Sci, 55(8):687-693.
[20] Large, M.J., DeMayo, F.J., 2012. The regulation of embryo implantation and endometrial decidualization by progesterone receptor signaling.
Mol Cell Endocrinol, 358(2):155-165.
[21] Lee, K.Y., Jeong, J.W., Wang, J., 2007. Bmp2 is critical for the murine uterine decidual response.
Mol Cell Biol, 27(15):5468-5478.
[22] Li, Q., Kannan, A., Wang, W., 2007. Bone morphogenetic protein 2 functions via a conserved signaling pathway involving Wnt4 to regulate uterine decidualization in the mouse and the human.
J Biol Chem, 282(43):31725-31732.
[23] Li, Y., Zhou, X., Wei, Q.W., 2014. Cell-specific expression and immunolocalization of nitric oxide synthase isoforms and soluble guanylyl cyclase α and β subunits in postnatal porcine uteri.
Acta Histochem, 116(3):466-473.
[24] Lin, H.Y., Wang, H.M., Li, Q.L., 2004. Expression of
Smad2 and
Smad4, transforming growth factor-β signal transducers in rat endometrium during the estrous cycle, pre-, and peri-implantation.
Anim Reprod Sci, 80(3-4):303-316.
[25] Liu, G., Lin, H., Zhang, X., 2004. Expression of
Smad2 and
Smad4 in mouse uterus during the oestrous cycle and early pregnancy.
Placenta, 25(6):530-537.
[26] Massague, J., Seoane, J., Wotton, D., 2005. Smad transcription factors.
Genes Dev, 19(23):2783-2810.
[27] Miyazono, K., Kamiya, Y., Morikawa, M., 2010. Bone morphogenetic protein receptors and signal transduction.
J Biochem, 147(1):35-51.
[28] Nagashima, T., Li, Q., Clementi, C., 2013. BMPR2 is required for postimplantation uterine function and pregnancy maintenance.
J Clin Invest, 123(6):2539-2550.
[29] Omwandho, C.O., Konrad, L., Halis, G., 2010. Role of TGF-βs in normal human endometrium and endometriosis.
Hum Reprod, 25(1):101-109.
[30] Pangas, S.A., 2012. Bone morphogenetic protein signaling transcription factor (SMAD) function in granulosa cells.
Mol Cell Endocrinol, 356(1-2):40-47.
[31] Paria, B.C., Ma, W.G., Tan, J., 2001. Cellular and molecular responses of the uterus to embryo implantation can be elicited by locally applied growth factors.
PNAS, 98(3):1047-1052.
[32] Shi, F., Stewart, R.L., Perez, E., 2004. Cell-specific expression and regulation of soluble guanylyl cyclase α
1 and β
1 subunits in the rat ovary.
Biol Reprod, 70(6):1552-1561.
[33] Shimasaki, S., Moore, R.K., Otsuka, F., 2004. The bone morphogenetic protein system in mammalian reproduction.
Endocr Rev, 25(1):72-101.
[34] Shimizu, T., Yokoo, M., Miyake, Y., 2004. Differential expression of bone morphogenetic protein 4–6 (BMP-4, -5, and -6) and growth differentiation factor-9 (GDF-9) during ovarian development in neonatal pigs.
Domest Anim Endocrinol, 27(4):397-405.
[35] Tanwar, P.S., McFarlane, J.R., 2011. Dynamic expression of bone morphogenetic protein 4 in reproductive organs of female mice.
Reproduction, 142(4):573-579.
[36] Wan, M., Cao, X., Wu, Y., 2002. Jab1 antagonizes TGF-β signaling by inducing Smad4 degradation.
EMBO Rep, 3(2):171-176.
[37] Wen, X., He, J., Zhang, X., 2011. Localization of Smad4 in the ovary of the European hedgehog (
Erinaceus europaeus L.).
Acta Histochem, 113(3):382-386.
[38] Wetendorf, M., DeMayo, F.J., 2012. The progesterone receptor regulates implantation, decidualization, and glandular development via a complex paracrine signaling network.
Mol Cell Endocrinol, 357(1-2):108-118.
[39] Xia, Y., Sidis, Y., Mukherjee, A., 2005. Localization and action of Dragon (repulsive guidance molecule b), a novel bone morphogenetic protein coreceptor, throughout the reproductive axis.
Endocrinology, 146(8):3614-3621.
[40] Yi, S.E., Lapolt, P.S., Yoon, B.S., 2001. The type I BMP receptor BmprIB is essential for female reproductive function.
PNAS, 98(14):7994-7999.
[41] Ying, Y., Zhao, G.Q., 2000. Detection of multiple bone morphogenetic protein messenger ribonucleic acids and their signal transducer, Smad1, during mouse decidualization.
Biol Reprod, 63(6):1781-1786.


Open peer comments: Debate/Discuss/Question/Opinion
<1>
Dr Romana<romzsarawr@gmail.com>
2017-08-23 14:45:19
Good article