Full Text:
![]() <3061>
<3061>
Summary: ![]() <2195>
<2195>
Suppl. Mater.: ![]()
CLC number: Q78
On-line Access: 2014-05-05
Received: 2013-12-31
Revision Accepted: 2014-04-04
Crosschecked: 2014-04-23
Cited: 6
Clicked: 5754
Early lethality of shRNA-transgenic pigs due to saturation of microRNA pathways
| ||||||||||||||
Zhen Dai, Rong Wu, Yi-cheng Zhao, Kan-kan Wang, Yong-ye Huang, Xin Yang, Zi-cong Xie, Chang-chun Tu, Hong-sheng Ouyang, Tie-dong Wang, Da-xin Pang. Early lethality of shRNA-transgenic pigs due to saturation of microRNA pathways[J]. Journal of Zhejiang University Science B, 2014, 15(5): 466-473.
@article{title="Early lethality of shRNA-transgenic pigs due to saturation of microRNA pathways",
author="Zhen Dai, Rong Wu, Yi-cheng Zhao, Kan-kan Wang, Yong-ye Huang, Xin Yang, Zi-cong Xie, Chang-chun Tu, Hong-sheng Ouyang, Tie-dong Wang, Da-xin Pang",
journal="Journal of Zhejiang University Science B",
volume="15",
number="5",
pages="466-473",
year="2014",
publisher="Zhejiang University Press & Springer",
doi="10.1631/jzus.B1400001"
}
%0 Journal Article
%T Early lethality of shRNA-transgenic pigs due to saturation of microRNA pathways
%A Zhen Dai
%A Rong Wu
%A Yi-cheng Zhao
%A Kan-kan Wang
%A Yong-ye Huang
%A Xin Yang
%A Zi-cong Xie
%A Chang-chun Tu
%A Hong-sheng Ouyang
%A Tie-dong Wang
%A Da-xin Pang
%J Journal of Zhejiang University SCIENCE B
%V 15
%N 5
%P 466-473
%@ 1673-1581
%D 2014
%I Zhejiang University Press & Springer
%DOI 10.1631/jzus.B1400001
TY - JOUR
T1 - Early lethality of shRNA-transgenic pigs due to saturation of microRNA pathways
A1 - Zhen Dai
A1 - Rong Wu
A1 - Yi-cheng Zhao
A1 - Kan-kan Wang
A1 - Yong-ye Huang
A1 - Xin Yang
A1 - Zi-cong Xie
A1 - Chang-chun Tu
A1 - Hong-sheng Ouyang
A1 - Tie-dong Wang
A1 - Da-xin Pang
J0 - Journal of Zhejiang University Science B
VL - 15
IS - 5
SP - 466
EP - 473
%@ 1673-1581
Y1 - 2014
PB - Zhejiang University Press & Springer
ER -
DOI - 10.1631/jzus.B1400001
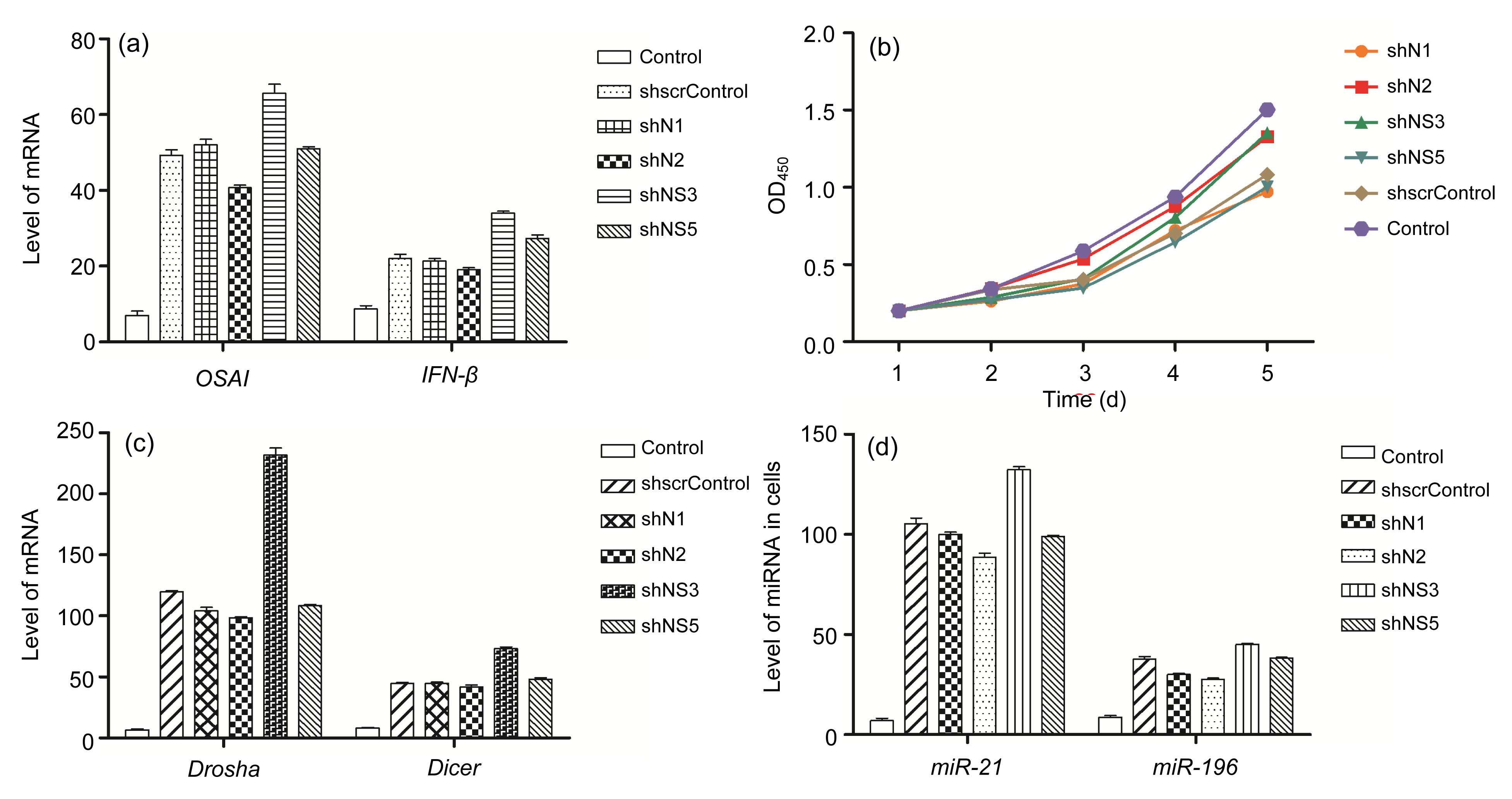
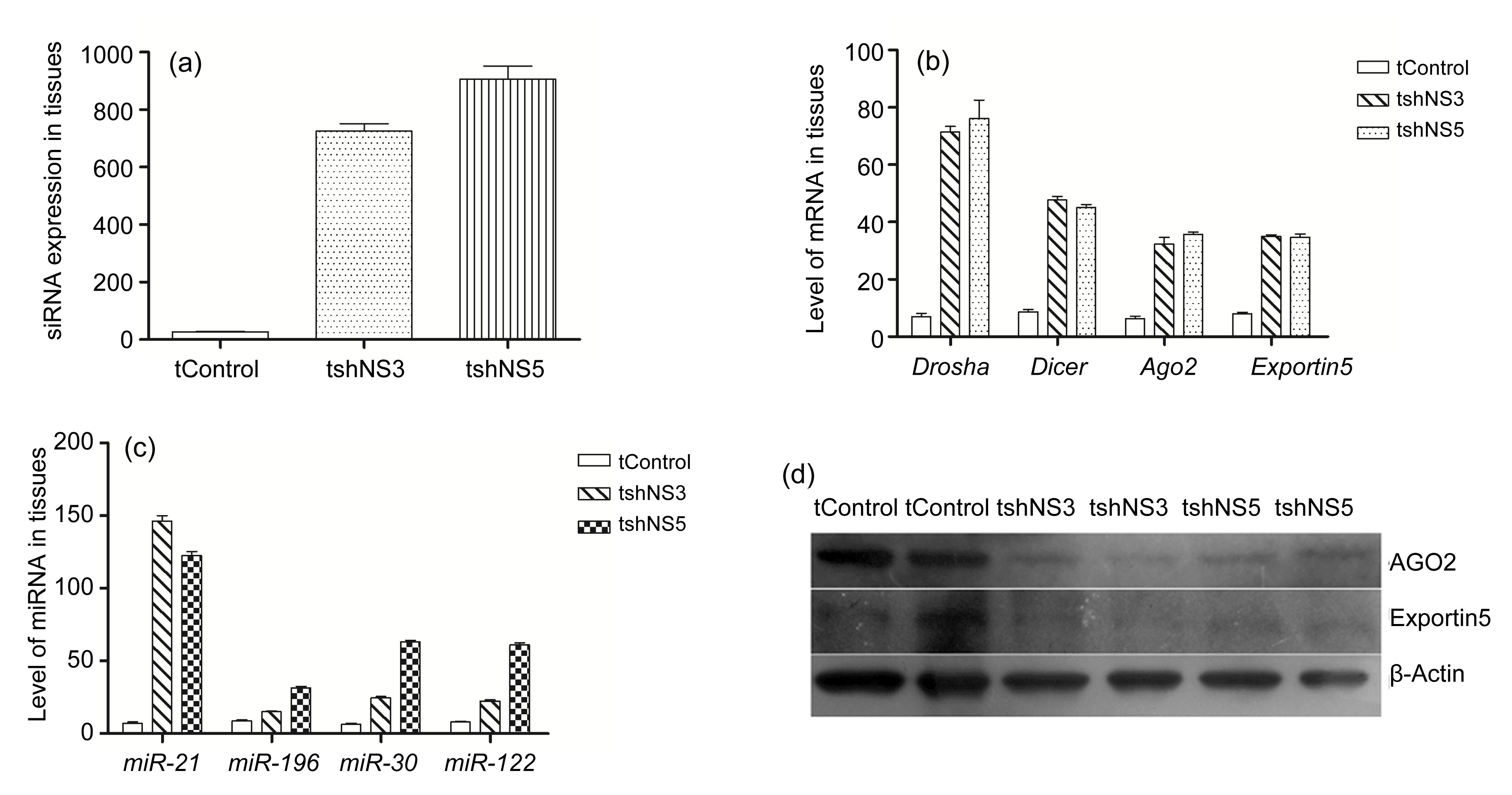
Abstract: RNA interference (RNAi) is considered as a potential modality for clinical treatment and anti-virus animal breeding. Here, we investigate the feasibility of inhibiting classical swine fever virus (CSFV) replication by short hairpin RNA (shRNA) in vitro and in vivo. We generate four different shRNA-positive clonal cells and two types of shRNA-transgenic pigs. CSFV could be effectively inhibited in shRNA-positive clonal cells and tail tip fibroblasts of shRNA-transgenic pigs. Unexpectedly, an early lethality due to shRNA is observed in these shRNA-transgenic pigs. With further research on shRNA-positive clonal cells and transgenic pigs, we report a great induction of interferon (IFN)-responsive genes in shRNA-positive clonal cells, altered levels of endogenous microRNAs (miRNA), and their processing enzymes in shRNA-positive cells. What is more, abnormal expressions of miRNAs and their processing enzymes are also observed in the livers of shRNA-transgenic pigs, indicating saturation of miRNA/shRNA pathways induced by shRNA. In addition, we investigate the effects of shRNAs on the development of somatic cell nuclear transfer (SCNT) embryos. These results show that shRNA causes adverse effects in vitro and in vivo and shRNA-induced disruption of the endogenous miRNA pathway may lead to the early lethality of shRNA-transgenic pigs. We firstly report abnormalities of the miRNA pathway in shRNA-transgenic animals, which may explain the early lethality of shRNA-transgenic pigs and has important implications for shRNA-transgenic animal preparation.
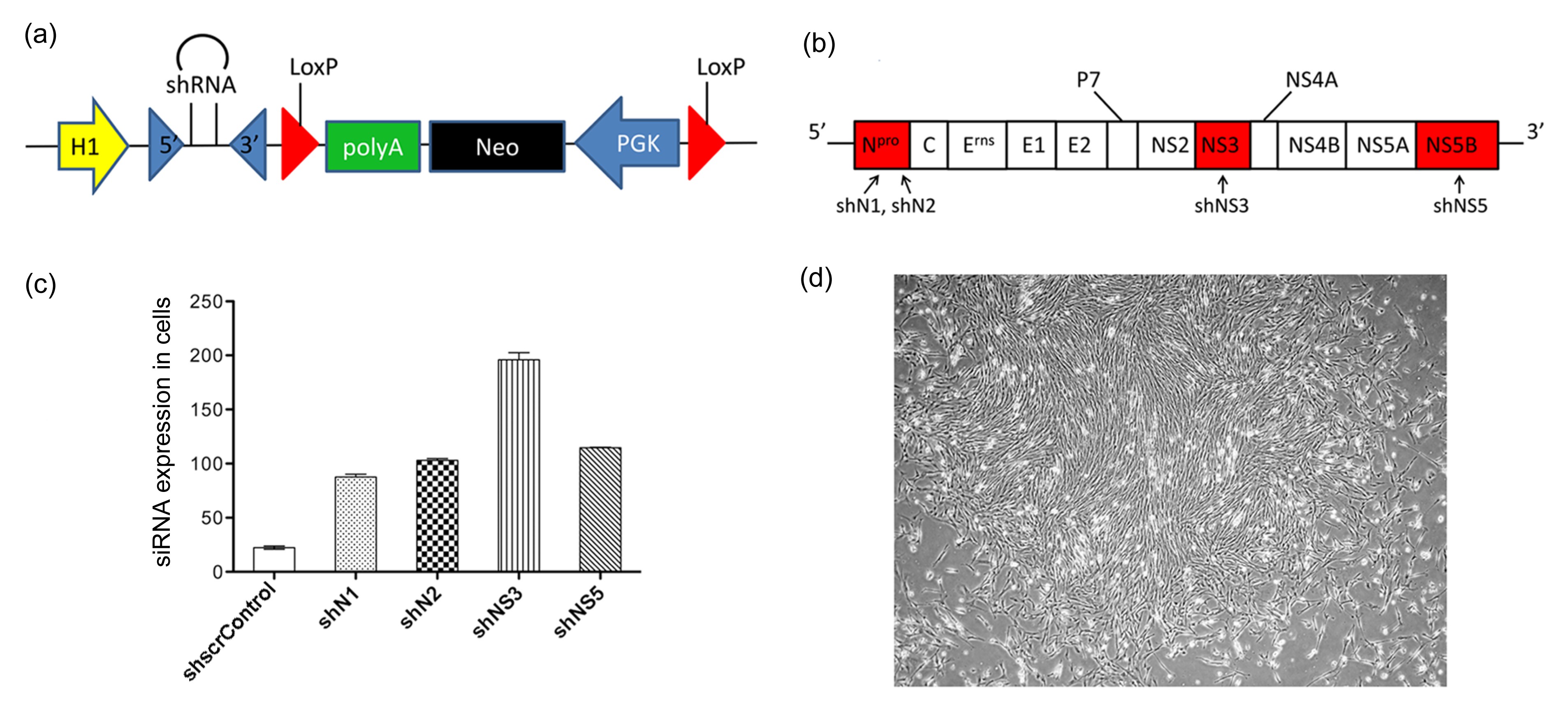
| Group | No. of embryos | No. (percent) of cleavages |
No. (percent) of blastocysts | |
| 24 h | 48 h | |||
| Control | 160 | 87 (54.38%)a | 140 (87.50%)a | 20 (12.50%)a |
| shscrControl | 162 | 85 (52.47%)a | 135 (83.33%)b | 20 (12.35%)a |
| shN1 | 148 | 90 (60.81%)b | 132 (89.19%)a | 16 (10.81%)a |
| shN2 | 152 | 80 (52.63%)a | 140 (92.10%)b | 20 (13.16%)a |
| shNS3 | 155 | 93 (60.00%)b | 137 (88.39%)a | 10 (6.45%)b |
| shNS5 | 158 | 84 (53.16%)a | 134 (84.81%)b | 13 (8.23%)b |



Open peer comments: Debate/Discuss/Question/Opinion
<1>