1. Introduction
IgA nephropathy (IgAN), characterized by immunoglobulin A depositing in the glomeruli (Berger and Hinglais,
1968; Woodroffe et al.,
1980; Lee et al.,
1982), is one of the major causes leading to renal failure (Cagnoli et al.,
1985; Imai et al.,
1985; Coppo et al.,
1986). In humans, telomere shortening takes place in many tissues with age, and is accelerated in chronic diseases (Jiang et al.,
2007). However, the contributing role of telomere dysfunction in human kidney disease was still unclear. Telomere length shortened during renal aging, especially in the cortex (Melk et al.,
2000). By activating the DNA damage responses, the telomere defect restricts the proliferative capacity of human cells, inducing senescence or apoptosis instead. Aging kidneys are significantly affected by the structural and functional changes, which leads to the reduction in glomerular filtration rate (GFR), a decline in renal hemodynamic and functional reserves, decreased sodium excretion and potassium clearance, diminished urinary concentration and diluting ability, susceptibility to drug toxicity, impaired endocrine function, and so on. Aging, being a complex process, involves a series of cross-linking molecular pathways. Understanding the molecular mechanism of the physiological or pathological aging process of kidneys could shed light on the clinical approaches to slow down renal aging and treat nephropathy. Several aging markers have been identified in renal aging (Halloran and Melk,
2001; Melk et al.,
2004). For convenient clinical use, more relevant biomarkers need to be detected not only in kidney tissue but also in urine and blood.
Previously, we have found that four proteins [cathelin-related antimicrobial peptide (CRAMP), stathmin, elongation factor-1α (EF-1α), and chitinase] were upregulated in telomere-dysfunctional mice. In geriatric population with aging-associated disorders and chronic diseases such as myelodysplastic syndrome (MDS) and liver cirrhosis, these proteins are positively regulated. However, there have been no reports on the relationship between the accumulation of these markers and the progression of IgAN. To further evaluate the conception that telomere attrition and aging process are interconnected to provoke human aging and disease, we investigated the association between these telomere shortening related markers and chronic kidney disease, IgAN, and we assessed the prognostic value of these markers for the progression of nephropathy in the renal biopsy, urine, and blood.
2. Materials and methods
2.1. Subjects
In this study, all IgAN patients (
n=177) and normal human (
n=83) controls provided blood, urine, and organ samples. The patients of IgAN are grouped depending on Lee’s grading system: IgAN I–II including grades I and II show normal or mild pathological changes such as focal or diffuse mesangial cell proliferation, and <25% of glomeruli with segmental sclerosis (SS)/global sclerosis (GS)/crescent (Cr); IgAN III+, which show obvious pathological changes and clinical disease progression, represent grades III, IV, and V, >25% of glomeruli with Cr/SS/GS, poorer kidney function, and large quantitative proteinuria. IgAN III+ patients have the worst kidney function compared with the controls and IgAN I–II. They show a higher concentration of serum creatinine, a lower modification of diet in renal disease (MDRD), and a higher proteinuria (Table
1). Healthy kidney tissues were obtained from 20 living kidney transplantation donors with written consent, and they show the longest telomere length in the three groups. There are 50 patients with systemic lupus erythematosus (SLE), and they were grouped depending on clinical symptoms and pathological changes: SLE I–III as the normal and mild groups; SLE IV–V as the progressed group. There were 30 patients with diabetic nephropathy (DN), and they were grouped depending on clinical symptoms and pathological changes into the early pathological change group or, with late pathological change, into the progressed group. There were 30 patients with focal segmental glomerulosclerosis (FSGS), and they were grouped depending on clinical symptoms and pathological changes into the early pathological change group or, with late pathological change, into the progressed group.
Table 1
Demographic and clinical data of the study cohorts
| Group |
n
|
Age (year) |
Gender (male/female) |
Serum creatine (μmol/L) |
Proteinuria (g/d) |
MDRD |
| Control |
83 |
35.3±9.9 |
44/39 |
70.6±12.6 |
|
96.8±15.6 |
| IgAN I–II |
74 |
34.0±10.9 |
35/39 |
74.5±16.3 |
1.0±1.0 |
95.9±21.8 |
| IgAN III+ |
103 |
36.0±13.7 |
52/50 |
124.0±91.0 |
1.9±1.3 |
49.7±18.5 |
| SLE I–III |
30 |
38.2±8.9 |
4/26 |
75.1±11.1 |
|
92.1±14.5 |
| SLE IV–V |
20 |
39.0±10.2 |
3/17 |
124.5±46.1 |
1.2±1.3 |
45.9±22.9 |
| DN early change |
20 |
35.2±11.2 |
9/11 |
74.0±22.0 |
|
99.7±12.9 |
| DN late change |
10 |
36.7±10.8 |
4/6 |
129.1±51.6 |
1.8±1.2 |
42.3±19.9 |
| FSGS early change |
20 |
37.1±9.5 |
9/11 |
78.5±19.6 |
|
90.3±18.2 |
| FSGS progressed change |
10 |
39.1±10.1 |
4/6 |
131.4±60.8 |
2.1±1.3 |
47.5±18.6 |
Data are expressed as mean±SD, except for gender
2.2. Direct enzyme-linked immunosorbent assay (ELISA)
The MicroWell plates (Nunc) were coated with the standard protein [CRAMP 100, 50, 25, 12.5, 6.25, 3.125, 1.6 and 0 ng, and 100 μl plasma (two-fold dilution)], and then incubated overnight at 4 °C. After three washes with phosphate buffered saline with Tween 20 (PBST), the plates were incubated with the first antibody [CRAMP (Cell Signaling) 1:100 (v/v), EF-1α (Upstate) 1:1000 (v/v), and stathmin (Cell Signaling Technology) 1:1000 (v/v)] for 2 h at ambient temperature or overnight at 4 °C, and then washed three times with PBST again. Horseradish peroxidase (HRP)-hapten conjugate secondary antibody 100 μl per well was applied, and the plates were incubated at room temperature for 60 min and washed five times with PBST. Then 0.1 ml of tetramethylbenzidine peroxide-based substrate solution was added to every well. The color reaction was developed for 30 min, and then stopped with the addition of 50 μl/well of 2 mol/L H
2SO
4 stop solution. The plate was read in the microplate reader in dual-wavelength mode (450–540 nm). The human CRAMP ELISA kit (HK321) we used is from Cell Sciences.
2.3. Chitinase activity
We analyzed chitinase activity with a chitinase assay kit (CS1030) purchased from Sigma. A 3-μl mixture of plasma and substrate solution (4-methylumbelliferone) was applied for each reaction. The enzyme activity was determined by using fluorimetry at an excitation wavelength of 360 nm and an emission wavelength of 450 nm. All samples were analyzed in triplicate.
2.4. Quantitative fluorescence in situ hybridization (qFISH)
qFISH was performed as described by Licastro et al. (
2003) on 5-μm thick paraffin sections of small intestine and methanol/acetic-acid fixed bone marrow cells, respectively. Telomere length was analyzed from the nuclei of the kidney cells using TFL-Telo Software from Peter Lansdorp.
2.5. Immunofluorescence staining for CRAMP
The expression of CRAMP was detected by immunofluorescence staining on 4-μm thick paraffin sections of kidney. After deparaffinization and rehydration, the sections were permeabilized in 1 mmol/L sodium citrate buffer for antigen retrieval. Primary antibody, CRAMP (1:100 (v/v) dilution; Cell Signaling), was applied for 2 h at room temperature or 4 °C overnight. The secondary antibody we used was anti-mouse Cy3-Zymed (1:200 (v/v)).
2.6. Statistical analysis
We used MedCalc software package (MedCalc for Windows 8.1.1.0), SPSS 16.0, and GraphPad Prism to perform statistical analysis. Normal distribution of the data was evaluated by the one-sample Kolmogorov-Smirnov test. Average values were presented with mean±standard deviation (SD) or median as appropriate.
P values were generated by the unpaired Student’s
t-test for all of the datasets.
3. Results
Healthy individuals (controls,
n=83) and patients with IgAN (
n=177) are matched in age and gender (Table
1). There are significant differences in serum creatine, proteinuria, and MDRD between IgAN I–II and IgAN III+ patients (
P values are 0.004, 0.032, and 0.004, respectively). Patients with SLE (
n=50), diabetic nephropathy (
n=30), and FSGS (
n=30) are all matched in age and gender (Table
1). There are also significant differences in serum creatine, proteinuria, and MDRD between SLE I–III and IV–V patients (
P=0.021), between DN early change and late change (
P=0.030), between FSGS early change and progressed change (
P=0.033). Data on blood and urine biochemistry at baseline were available for all healthy individuals, and data on kidney histology were obtained from 20 living kidney transplantation donors with written consent.
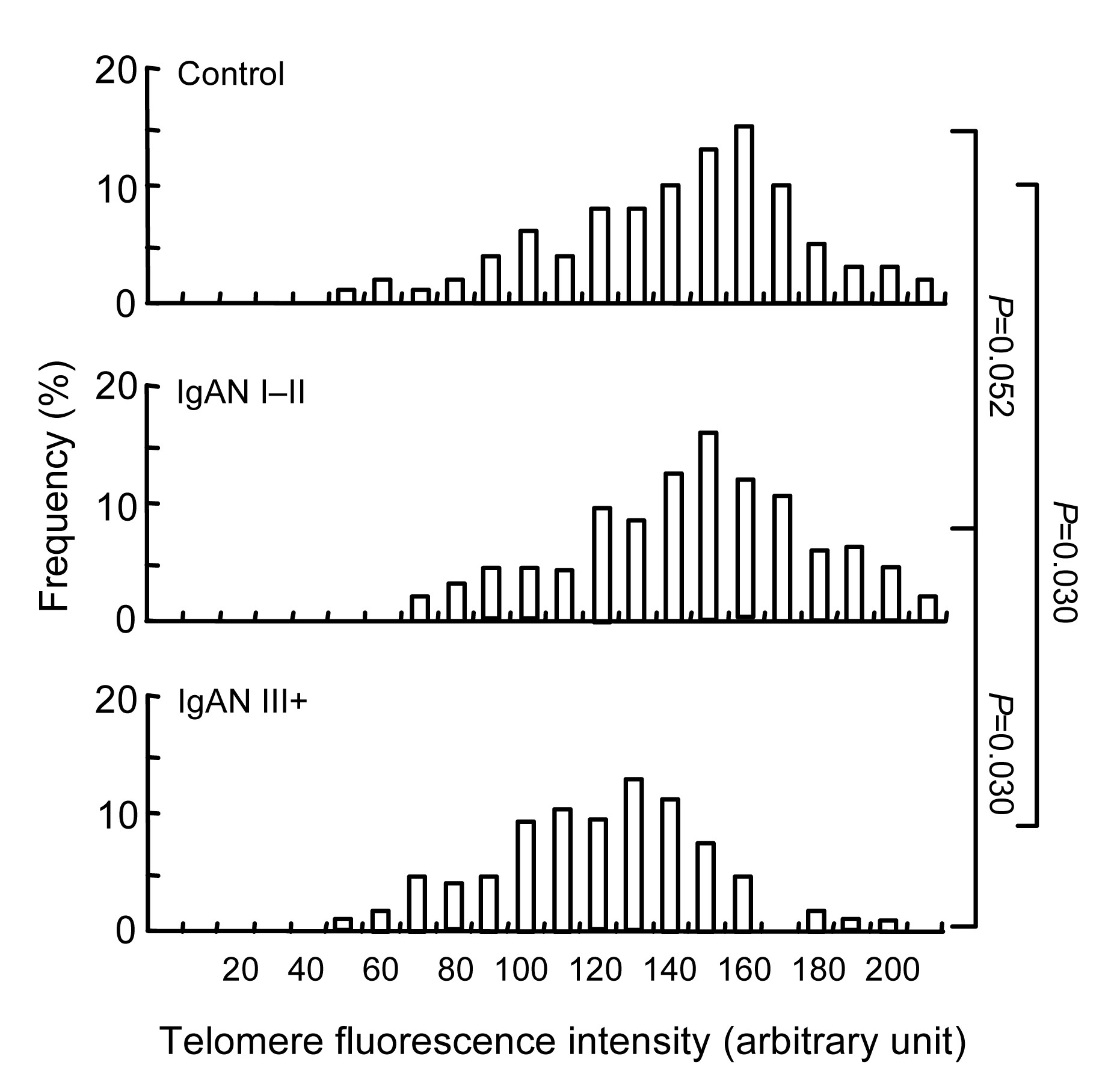
3.1. Telomere length in different patients
Telomere length was determined in all IgAN, SLE, DN, and FSGS patients. Data on kidney histology were available from all patients, and data on blood and urine biochemistry at the baseline (time of biopsy) were available from all patients. The patients with IgAN III+ have the shortest telomere length compared with the controls (
P=0.030; Fig.
1) and the IgAN I–II group. Patients with IgAN I–II have a shorter telomere length compared with the healthy controls, although the
P value does not reach a statistical significance (
P=0.052; Fig.
1). Telomere length has no significant difference between patients with SLE I–III and IV–V, between patients with early DN and the late pathological change group, between patients with early FSGS and the progression group.
Fig.1
Distribution of telomere length (telomere fluorescence intensity) in kidney cells from the healthy controls, IgAN I–II and IgAN III+ patients
Bars to the left side of the vertical line represent cells with critically short telomeres; the percentage of such cells is indicated
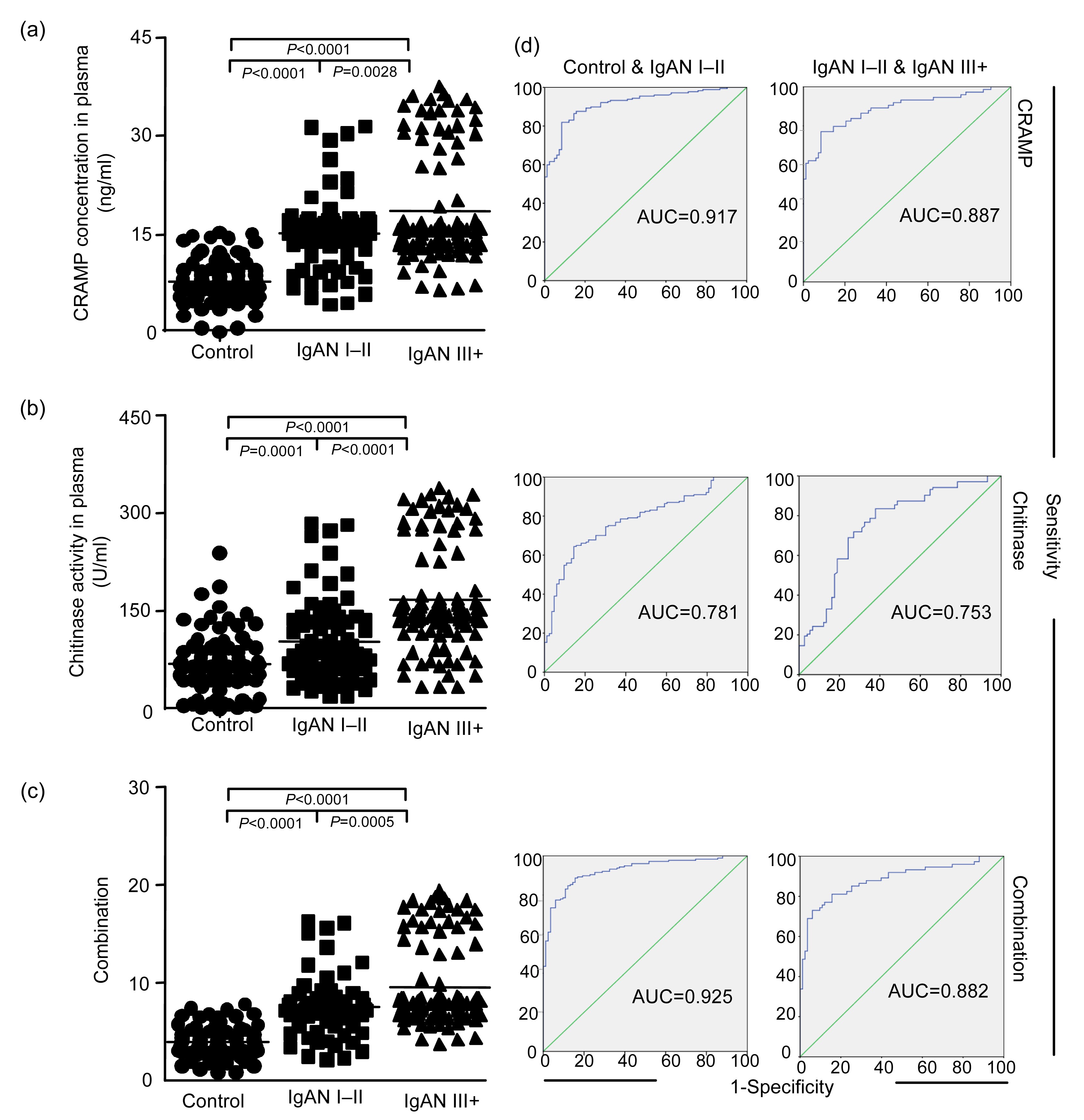
3.2. Expression levels of biomarkers in blood plasma
ELISA is used to detect plasma samples and urine samples from all subjects, and internal standards are applied to normalize the data. In blood plasma, the expression level of CRAMP (
P<0.0001; Fig.
2a) and the level of chitinase activity (
P=0.0001; Fig.
2b) are higher in the patients with IgAN I–II than in the controls, whereas the differences in EF-1α and stathmin expression levels are not significant between these two groups. There is a further increase in plasma levels of CRAMP (
P=0.0028) and chitinase activity (
P<0.0001) in the patients with IgAN III+ compared with the IgAN I–II group. Multivariate analysis indicated that the expression level of CRAMP (
A) and chitinase activity (
B) have the best discriminatory power (combination score=0.434
A+0.009
B; Table
S1) and the combination of these two markers has the highest statistical power to discriminate between the three groups (Figs. 2c and 2d). Plasma protein expression shows no significant difference between the patients with SLE I–III and IV–V, between the patients in early DN and late pathological change groups, between the patients in early FSGS and progressed pathological change groups.
Fig.2
Increased expression of proteins induced by telomere dysfunction in plasma during human IgAN progression (a–c) and the specificity and sensitivity of the marker proteins using receiver operating characteristic (ROC) analysis and comparing the healthy group with IgAN groups (d)
The dot plots show the levels of the indicated marker proteins in human blood plasma as measured by ELISA for CRAMP concentration (a), chitinase activity (b) (note that CRAMP concentration and chitinase activity were up-regulated in the mild and moderate IgAN groups step by step compared with the healthy group), and a combined analysis (c) of the best two markers (CRAMP and chitinase) according to the regression analysis (Table S1) on the same set of sample
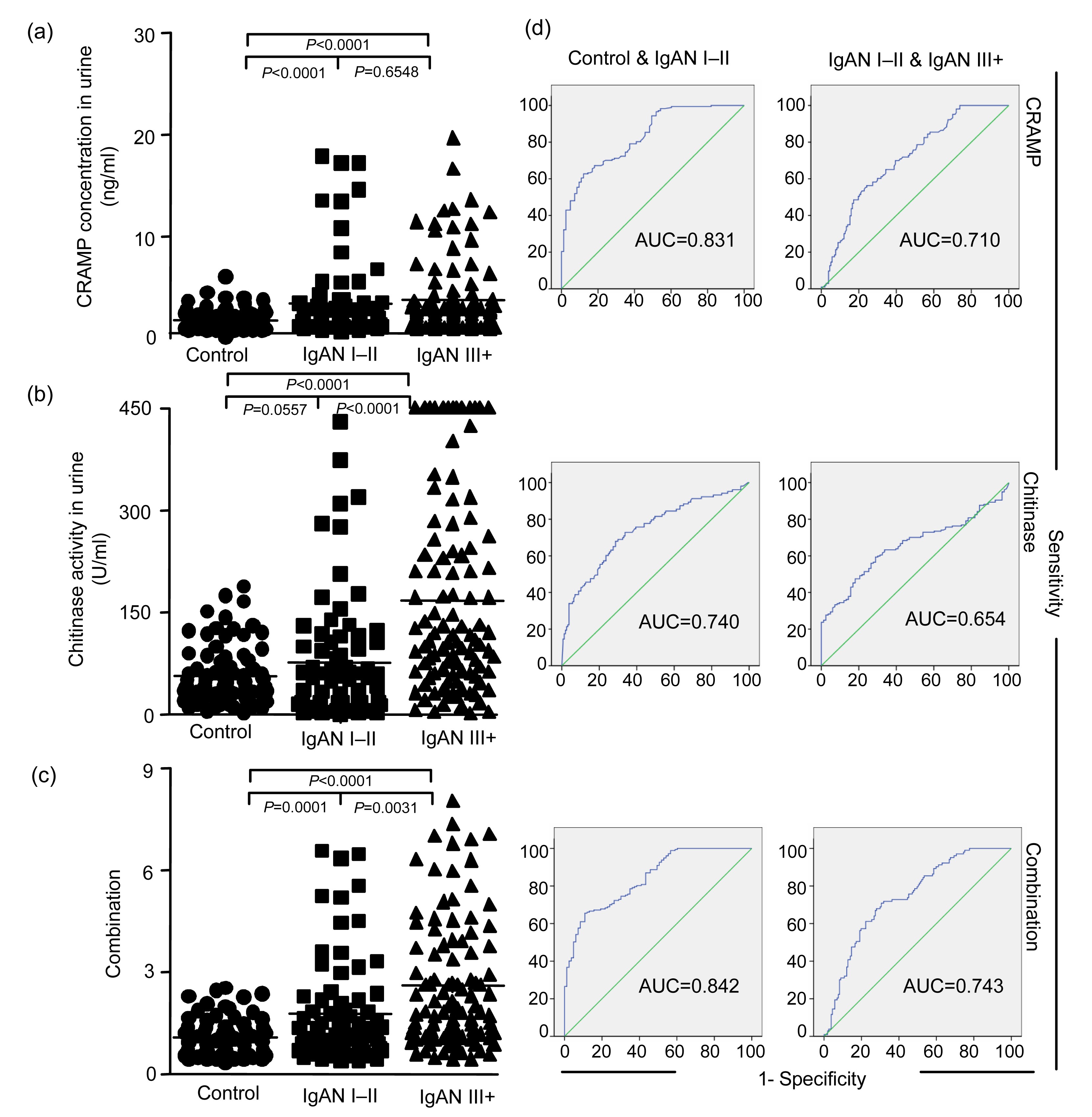
3.3. Expression levels of biomarkers in urine
In urine, the expression level of CRAMP (P<0.0001; Fig. 3a) and the level of chitinase activity (P=0.0049; Fig. 3b) are higher in the patients with IgAN I–II than in controls, whereas the differences in EF-1α and stathmin expression levels are not significant between these two groups. There is a further increase in the urine level of chitinase activity (P<0.0001) in the patients with IgAN III+. However, for CRAMP (P=0.6548), EF-1α, and stathmin, the differences are not significant between the patients with IgAN I–II and IgAN III+. Multivariate analysis indicated that the expression level of CRAMP (A) and chitinase activity (B) have the best discriminatory power (combination score=0.326A+0.008B; Table S2) and a combination of these two markers has the highest statistical power to discriminate between the three groups (Figs. 3c and 3d). There is no statistical difference in urine protein expression between the patients with SLE I–III and IV–V, between the patients in early DN and the late pathological change groups, between the patients in early FSGS and the progression groups.
Fig.3
Increased expression of proteins induced by telomere dysfunction in urine during human IgAN progression (a–c) and the specificity and sensitivity of the marker proteins using receiver operating characteristic (ROC) analysis and comparing the healthy group with IgAN groups (d)
The dot plots show the levels of the indicated marker proteins in human urine as measured by ELISA for CRAMP concentration (a), chitinase activity (b) (note that the two marker proteins were up-regulated in the mild and moderate IgAN groups step by step compared with healthy group), and a combined analysis (c) of the best two markers (CRAMP and chitinase) according to the regression analysis (Table S2) on the same set of sample
3.4. Expression levels of CRAMP protein by immunofluorescence staining
Immunofluorescence staining confirmed the up-regulation of CRAMP protein levels in the patients with IgAN III+ (Fig. 4). The percentage of CRAMP positive cells in IgAN III+ patients is higher than that in IgAN I–II patients (P=0.031) and controls (P=0.016).
Fig.4
Increased expression of biomarkers in the kidney of IgAN patients
(a) Representative immunohistochemistry staining of CRAMP in post mortem kidney biopsies of unaffected, healthy controls (live doner of kidney transplantation, n=20) compared with unaffected, IgAN I–II and IgAN III+ patients; (b) The percentage of organ cells that are positive staining for CRAMP in the kidneys
3.5. Relationship between the expression levels of biomarkers and MDRD
In blood plasma and urine, the expression levels of CRAMP, EF-1α, and stathmin, and chitinase activity (Figs. S1 and S2) do not correlate with MDRD. The combination scores of CRAMP and chitinase activity also do not correlate with MDRD (Figs. S1e and S2e).
4. Discussion
In this study, we have found that telomere shortening and aging markers are associated with the progression of IgAN, and not associated with the progression of SLE, DN, or FSGS. Previously, we have found that these biomarkers increased in humans with liver cirrhosis or myelodysplatic syndrome diseases, which also proved to be associated with telomere shortening (Ohyashiki et al., 1994; Wiemann et al., 2002). Telomere dysfunction provokes defects in the microenvironment and macroenvironment in vivo, by increasing the expression of various cytokines such as granulocyte colony-stimulating factor (G-CSF) in the plasma. As a result, the function of bone marrow stromal cells is impaired and functional hematopoietic stem cells are reduced as well (Ju et al., 2007). It is reported that telomeres appear to be especially sensitive to chronic oxidative stress, since telomere length is decreasing at a faster speed during the accumulation of reactive oxygen species (ROS) induced 8-oxodG DNA-strand breaks (Houben et al., 2008). Therefore, evaluation of telomere length might be an effective biomarker for disease progression (Houben et al., 2008).
Since the renal function can be evaluated relatively easily in clinical practice and has been quantified in longitudinal studies, it is no surprise that kidney aging has become a general model for organ aging (Lindeman et al., 1985). The phenotype of human renal senescence can represent a phenotype of mass, particularly in the cortex (Takubo et al., 2002). There are some reports that have examined the relationship between kidney aging and telomere dysfunction (Lindeman et al., 1985; Lindeman, 1986; Gourtsoyiannis et al., 1990; Takubo et al., 2002; von Schnakenburg et al., 2002), and chronic renal disease is also influenced by telomeres triggering aging (Melk et al., 2003; Tchakmakjian et al., 2004; Szeto et al., 2005).
In IgAN patients, the combination scores of these four markers in the plasma and urine were up-regulated in the IgAN I–II patients and further up-regulated in the IgAN III+ patients, which were linked to telomere attrition. Probably, telomere shortening induces alterations in the function of cells and organs by up-regulating the expression of secreted proteins during human aging and chronic disease. Szeto et al. (2005) showed that the chromosomal telomere of kidney cells shortened in IgAN by the measurement of DNA in the urinary sediment. The overexpression of the aging marker p16 in the aging renal and chronic kidney disease has been reported to be correlated with telomere dysfunction (Melk et al., 2003; Szeto et al., 2005; Jiang et al., 2007; Zhou et al., 2008). We also found that the discrimination power of these markers measured in plasma was better than that measured in urine samples. One of the reasons could be that the concentrations of these proteins were much lower in urine than in plasma. It is also reported that there is a loss of renal function, characterized by increased renovascular resistance, reduced effective renal plasma flow (RPF), and decreased filtration fraction (Lindeman et al., 1985; Lindeman, 1986; Melk et al., 2005). However, in our current study, we could not find the correlation between these marker proteins and the renal function. We have found a trend for CRAMP/chitinase correlating with MDRD. Larger cohorts and longer follow-up clinical research need to be designed to fully address this question.
CRAMP and chitinase are active during innate immune responses in inflammatory diseases such as asthma, allergy, and parasitic infections (Fliser et al., 1993; Nizet et al., 2001). The activation of innate immunity potentially triggers the onset of decreased longevity and age-related inflammatory diseases like atherosclerosis (Reese et al., 2007; Vasto et al., 2007). Some studies suggest that telomere depletion involves the activation of innate immunity during human aging and chronic disease. The expression levels of these markers were significantly high in kidney tissues, and the positive staining was positioned in the renal interstitium. That is also combined with the pathological phenotype of serious IgAN with interstitial fibrosis and inflammatory cell infiltration. We hypothesized that DNA damage and telomere shortening triggered a higher expression of CRAMP and CRAMP stimulated the inflammatory cell infiltration, and that these could be correlated with renal disease, and redesigned the project to find something interesting.
The degree of DNA damage and telomere shortening varies in different diseases. In this paper, we found that the difference in telomere shortening was not significant in FSGS, SLE, or DN progression, probably because the DNA damage was not serious enough for qFISH to catch. It means that the mechanisms of SLE (immune system disorder), DN (glucose metabolic dysfunction), and FSGS (podocytopathy with reduction in podocytes density) diseases are different from that of IgAN.
In summary, the results reveal that IgAN is associated with the increase in secreted proteins due to telomere attrition. These findings are in consistence with previous studies respecting telomere dysfunction in kidney aging and nephropathy (Melk et al., 2003; Tchakmakjian et al., 2004; Szeto et al., 2005; Jiang et al., 2007; Zhou et al., 2008). The recently identified biomarkers can be described as an effective indicator of human IgAN progression correlated with telomere dysfunction.
* Project supported by the National Basic Research Program (973) of China (Nos. 2011CB944002 and 2012CB517603), the National Natural Science Foundation of China (No. 2011BAI10B07), and the Major Special Project of Technology Office in Zhejiang Province (No. 2012C13G2010133), China# Electronic supplementary materials: The online version of this article (http://dx.doi.org/10.1631/jzus.B1300115) contains supplementary materials, which are available to authorized usersCompliance with ethics guidelines Ying-ying LU, Xian YANG, Wen-qing CHEN, Zhen-yu JU, Zhang-fei SHOU, Juan JIN, Xiao-hui ZHANG, Jiang-hua CHEN, and Hong JIANG declare that they have no conflict of interest.
References
[1] Berger, J., Hinglais, N., 1968. Intercapillary deposits of IgA-IgG.
J Urol Nephrol, 74(9):694-695.
[2] Cagnoli, L., Beltrandi, E., Pasquali, S., 1985. B and T cell abnormalities in patients with primary IgA nephropathy.
Kidney Int, 28(4):646-651.


[3] Coppo, R., Basolo, B., Rollino, C., 1986. Dietary gluten and primary IgA nephropathy.
N Engl J Med, 315(18):1167-1168.
[4] Fliser, D., Zeier, M., Nowack, R., 1993. Renal functional reserve in healthy elderly subjects.
J Am Soc Nephrol, 3(7):1371-1377.
[5] Gourtsoyiannis, N., Prassopoulos, P., Cavouras, D., 1990. The thickness of the renal parenchyma decreases with age: a CT study of 360 patients.
AJR Am J Roentgenol, 155(3):541-544.
[6] Halloran, P.F., Melk, A., 2001. Renal senescence, cellular senescence, and their relevance to nephrology and transplantation.
Adv Nephrol Necker Hosp, 31:273-283.
[7] Houben, J.M., Moonen, H.J., van Schooten, F.J., 2008. Telomere length assessment: biomarker of chronic oxidative stress?.
Free Radic Biol Med, 44(3):235-246.
[8] Imai, H., Nakamoto, Y., Asakura, K., 1985. Spontaneous glomerular IgA deposition in ddY mice: an animal model of IgA nephritis.
Kidney Int, 27(5):756-761.
[9] Jiang, H., Ju, Z., Rudolph, K.L., 2007. Telomere shortening and ageing.
Z Gerontol Geriatr, 40(5):314-324.
[10] Ju, Z., Jiang, H., Jaworski, M., 2007. Telomere dysfunction induces environmental alterations limiting hematopoietic stem cell function and engraftment.
Nat Med, 13(6):742-747.
[11] Lee, S.M., Rao, V.M., Franklin, W.A., 1982. IgA nephropathy: morphologic predictors of progressive renal disease.
Hum Pathol, 13(4):314-322.
[12] Licastro, F., Grimaldi, L.M., Bonaf, M., 2003. Interleukin-6 gene alleles affect the risk of Alzheimer’s disease and levels of the cytokine in blood and brain.
Neurobiol Aging, 24(7):921-926.
[13] Lindeman, R.D., 1986. The aging kidney.
Compr Ther, 12(3):43-49.
[14] Lindeman, R.D., Tobin, J., Shock, N.W., 1985. Longitudinal studies on the rate of decline in renal function with age.
J Am Geriatr Soc, 33(4):278-285.
[15] Melk, A., Ramassar, V., Helms, L.M., 2000. Telomere shortening in kidneys with age.
J Am Soc Nephrol, 11(3):444-453.
[16] Melk, A., Kittikowit, W., Sandhu, I., 2003. Cell senescence in rat kidneys
in vivo increases with growth and age despite lack of telomere shortening.
Kidney Int, 63(6):2134-2143.
[17] Melk, A., Schmidt, B.M., Takeuchi, O., 2004. Expression of p16
INK4a and other cell cycle regulator and senescence associated genes in aging human kidney.
Kidney Int, 65(2):510-520.
[18] Melk, A., Schmidt, B.M., Vongwiwatana, A., 2005. Increased expression of senescence-associated cell cycle inhibitor p16
INK4a in deteriorating renal transplants and diseased native kidney.
Am J Transplant, 5(6):1375-1382.
[19] Nizet, V., Ohtake, T., Lauth, X., 2001. Innate antimicrobial peptide protects the skin from invasive bacterial infection.
Nature, 414(6862):454-457.
[20] Ohyashiki, J.H., Ohyashiki, K., Fujimura, T., 1994. Telomere shortening associated with disease evolution patterns in myelodysplastic syndromes.
Cancer Res, 54(13):3557-3560.
[21] Reese, T.A., Liang, H.E., Tager, A.M., 2007. Chitin induces accumulation in tissue of innate immune cells associated with allergy.
Nature, 447(7140):92-96.
[22] Szeto, C.C., Poon, P.Y., Lai, F.M., 2005. Chromosomal telomere shortening of kidney cells in IgA nephropathy by the measurement of DNA in urinary sediment.
Clin Nephrol, 64(5):337-342.
[23] Takubo, K., Izumiyama-Shimomura, N., Honma, N., 2002. Telomere lengths are characteristic in each human individual.
Exp Gerontol, 37(4):523-531.
[24] Tchakmakjian, L., Gardner, J.P., Wilson, P.D., 2004. Age-dependent telomere attrition as a potential indicator of racial differences in renal growth patterns.
Nephron Exp Nephrol, 98(3):e82-e88.
[25] Vasto, S., Candore, G., Balistreri, C.R., 2007. Inflammatory networks in ageing, age-related diseases and longevity.
Mech Ageing Dev, 128(1):83-91.
[26] von Schnakenburg, C., Strehlau, J., Ehrich, J.H.H., 2002. Quantitative gene expression of TGF-β1, IL-10, TNF-α and Fas Ligand in renal cortex and medulla.
Nephrol Dial Transplant, 17(4):573-579.
[27] Wiemann, S.U., Satyanarayana, A., Tsahuridu, M., 2002. Hepatocyte telomere shortening and senescence are general markers of human liver cirrhosis.
FASEB J, 16(9):935-942.
[28] Woodroffe, A.J., Gormly, A.A., McKenzie, P.E., 1980. Immunologic studies in IgA nephropathy.
Kidney Int, 18(3):366-374.
[29] Zhou, X.J., Rakheja, D., Yu, X., 2008. The aging kidney.
Kidney Int, 74(6):710-712.


Open peer comments: Debate/Discuss/Question/Opinion
<1>