1. Introduction
There is a close relationship between malignant ventricular arrhythmias and myocardial infarction (MI) (Haugaa et al.,
2010), which leads to high cardiovascular mortality (Wang,
2012). It is well recognized that mechanical restitution and regional inhomogeneity in contractility act as foci for arrhythmias (Thygesen and Uretsky,
2004; Ashikaga et al.,
2005) through a process known as mechanoelectric feedback (MEF). In chronic MI, scar tissue is surrounded by normal or ischemic myocardium, and greater stretch occurs than in normal myocardium (Bertini et al.,
2010). Previous studies also verified that MEF can be increased by chronic MI (Kiseleva et al.,
2000), contributing to the appearance of arrhythmias. We inferred that electrophysiological changes induced by stretch in normal hearts, shortened action potential duration (APD), the occurrence of after-depolarization, and premature beats (Kiseleva et al.,
2000; Ravens,
2003; Taggart and Lab,
2008) might be observed in chronically infarcted myocardium. Many studies have demonstrated that stretch-activated ion channels (SACs) have a crucial role to perform in MEF, though details of the mechanism are unclear (Garan et al.,
2005; Lab,
2006).
Streptomycin (SM), a blocker of SACs, has been known to inhibit the activities of SACs in single cell, tissue, and intact preparations in various species (Lab,
2006). It can decrease the occurrence of arrhythmias induced by changes in ventricular wall-stress in isolated rat hearts (Salmon et al.,
1997). Eckardt et al. (
2001) studied Langendorff-perfused rabbit hearts and found that SM (200 μmol/L) could inhibit the shortened repolarization induced by stretching. The dominant frequency of ventricular fibrillation (VF) induced by stretching decreases following SM treatment in rabbit hearts (Trapero et al.,
2008). SM also prevents stretch-related vulnerability to atrial fibrillation (Ninio and Saint,
2008). In hearts from hypertensive rats, Kim et al. (
2012) recently found that 100 μmol/L SM could reduce stretch-induced ectopic heartbeats by increasing the threshold for stretch-induced arrhythmias.
However, the effect of blocking of SACs with SM has seldom been observed
in vivo, and results are often contradictory. Cooper and Kohl (
2005) reported that SM could not block chronotropy in the spontaneous pacemaking rate induced by stretching, and further questioned the
in situ utility of SM for acute blocking of mechanosensitive mechanisms. Garan et al. (
2005) found that SM reduced ST segment elevation following impact in swine but did not reduce the frequency of VF. Using skeletal muscle, Yeung et al. (
2005) and Spangenburg and McBride (
2006) found that oral SM
in vivo could block the activities of SACs in the rat and mouse, respectively. However, there has been no analogous study on myocardium.
The aim of this study was to observe the effect of stretching on the monophasic action potential (MAP) and the effect of the application of SM
in vivo on electrophysiological changes in chronically infarcted rat hearts.
2. Materials and methods
2.1. Preparation of MI model
The study was performed in accordance with the rules of
The Care and Use of Laboratory Animals issued by the US National Institute of Health (NIH Publication No. 85-23, revised in 1985).
Sixty Wistar rats (regardless of their genders, 180–224 g) were chosen for the experiment. Each rat, anaesthetized by intraperitoneal injection using sodium pentobarbital (40–50 mg/kg) and heparin (1000 U/kg), was intubated and then connected to a respirator. Artificial respiration was provided during the whole surgical process (breathing rate 60 min
−1, tidal volume 30 ml/100 mg). Electrocardiograms (ECG; limb leads including I, II, III, aVR, aVL, aVF) were recorded from electrodes attached to the limbs of each rat. Briefly, MI was obtained by ligating the left anterior descending branch (LAD) of the coronary artery. First, the heart was exteriorized between the 3rd and 4th costal bones of the left sternal border. Second, the LAD was ligated between the pulmonary conus and the left auricle. Successful ligature of the LAD was verified by regional myocardial cyanosis and ST elevation in the ECG. Third, the heart was replaced and the cut was stitched. Sham-operated animals were operated following the procedure mentioned above, but without ligation of the LAD. The surviving rats were kept in a cage and fed with a full diet and water.
Sham-operated animals were divided at random into two groups: a control group (
n=15) in which experiments were carried out after 8 weeks and an SM group (
n=15) in which after 7 weeks, the rats were injected intramuscularly with SM (Merro Pharmaceuticals Companies; 180 mg/(kg·d)) for 7 d. Experiments were then conducted. MI animals were also divided into two groups: an MI group (
n=15) in which experiments were carried out 8 weeks after ligature of the LAD and an MI+SM group (
n=15) in which, after ligation of the LAD for 7 weeks, the rats were injected intramuscularly with SM (180 mg/(kg·d)) for 7 d. Experiments were then conducted.
2.2. Recording
Each animal was anaesthetized and ventilated as mentioned above. ECG was recorded during the whole experiment. The right common carotid artery was dissociated. A catheter was connected to a pressure transducer (YP500, Gaobeidian, China), which was inserted into the left ventricle (LV) via the right common carotid artery and aorta by catheterization to measure the pressure of the ventricle. The distance between the end of the catheter and the transducer was minimized to decrease system dampening. Thoracotomy was performed. The heart and aortic arch were revealed. A suction electrode, placed on the anterior wall of the LV, was used to detect LV MAP (Fu et al.,
2007).
2.3. Protocols
After stabilization for 15 min, the heart was stretched by an occlusion of the aortic arch for 5 s, followed by unclamping of the aortic arch. The 90% monophasic action potential duration (MAPD
90), LV pressure, premature ventricular beats (PVBs), and ventricular tachycardia (VT) were monitored during 5 s of stretching.
2.4. Data analysis
The measurement and analysis of MAPD
90 (in ms) were conducted according to the criteria of Fu et al. (
2007). Every five beats, one MAP was analyzed and then averages were calculated. Stretch-induced arrhythmias (SIAs) were defined according to the guidelines of the Lambeth Convention (Curtis et al.,
2013). Data were analyzed using SPSS 13.0 (Statistical Package for the Social Sciences 13.0, SPSS Company, USA) by an independent University-based statistician. Quantitative data, expressed as mean±standard deviation (SD), were compared with analysis of variance (ANOVA). A
χ
2 test was used for qualitative data.
P<0.05 was considered statistically significant.
3. Results
3.1. Effect of stretching on MAPD90
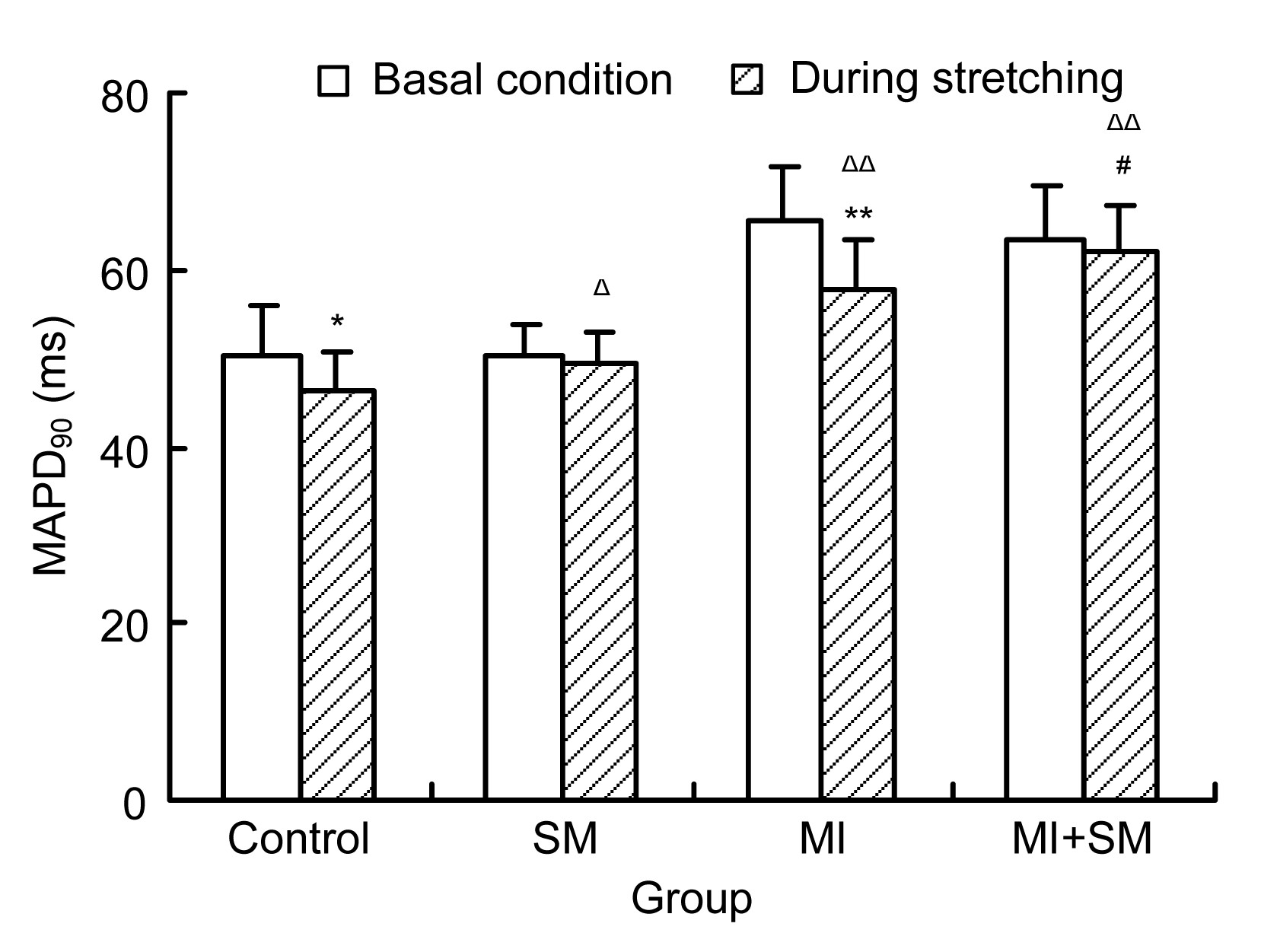
During the occlusion of the aortic arch, the MAPD
90 decreased, compared with the basal condition in the control group ((50.27±5.61) ms vs. (46.27±4.51) ms,
P<0.05) and in the MI group ((65.47±6.38) ms vs. (57.47±5.76) ms,
P<0.01) (Fig.
1). The MAPD
90 increased significantly compared with the control groups after ligation of the LAD ((65.47±6.38) ms vs. (50.27±5.61) ms,
P<0.01) and during stretching ((57.47±5.76) ms vs. (46.27±4.51) ms,
P<0.01) (Fig.
1).
Fig.1
Influence of streptomycin (SM) on 90% monophasic action potential duration (MAPD90) in control and myocardial infarction (MI) groups during stretching (the occlusion of the aortic arch for 5 s)
MAPD90 decreased in the control group during stretching (*
P<0.05) and in the MI group (**
P<0.01), compared with that under basic conditions. SM had no influence on MAPD90 in either normal or infarcted rat hearts under basic conditions, but SM did inhibit the decrease in MAPD90 induced by stretching (Δ
P<0.05, ΔΔ
P<0.01, compared with the control group; #
P<0.05, compared with the MI group). Data are expressed as mean±SD (n=15)
3.2. Effect of SM on MAPD90 changes during stretching
After the treatment with SM, the MAPD
90 showed no significant changes from basal conditions either in normal ((50.27±5.61) ms vs. (50.33±3.62) ms,
P>0.05) or infarcted rat hearts ((65.47±6.38) ms vs. (63.20±6.32) ms,
P>0.05) (Fig.
1). However, SM inhibited the stretch-induced decrease in MAPD
90 during the occlusion of the aortic arch, both in the normal (compared with the basal conditions, (49.53±3.52) ms vs. (50.33±3.62) ms,
P>0.05) and MI hearts (compared with the basal conditions, (61.87±5.33) ms vs. (63.20±6.32) ms,
P>0.05). SM partly reversed the decrease in MAPD
90 induced by stretching in the normal hearts ((49.53±3.52) ms vs. (46.27±4.51) ms,
P<0.05) and in the infarcted rat hearts ((61.87±5.33) ms vs. (57.47±5.76) ms,
P<0.05) (Fig.
1).
3.3. Effect of SM on SIAs
The PVBs appeared during the occlusion of the aorta. The number of PVBs was counted during the 5 s of occlusion of the aorta. The incidence of PVBs in the control group was observed during stretching (1.80±0.86). After ligation of the LAD for eight weeks, the incidence showed a marked and significant increase (7.93±1.66 in the MI group vs. 1.80±0.86 in the control group,
P<0.01). SM obviously reduced the number of PVBs caused by the occlusion of the aorta (1.80±0.86 vs. 0.93±0.59 in normal hearts,
P<0.05; 7.93±1.66 vs. 5.40±1.18 in infarcted hearts,
P<0.01) (Table
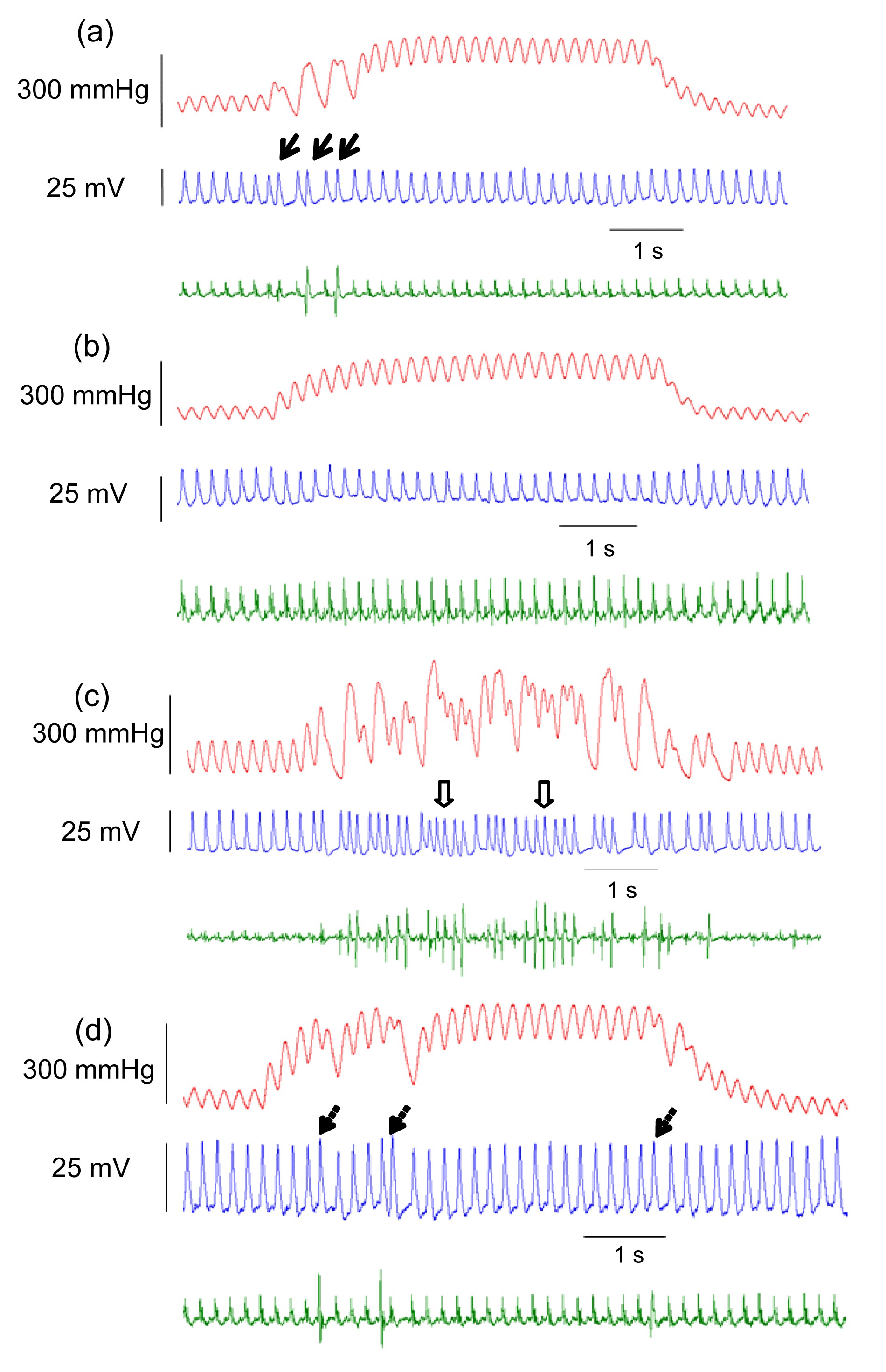
1). Signal collecting systems recorded the MAP figures (Fig.
2).
Table 1
Effect of streptomycin (SM) on premature ventricular beats (PVBs) induced by stretching for 5 s
| Group |
Number of PVBs |
| Control |
1.80±0.86 |
| SM |
0.93±0.59*
|
| MI |
7.93±1.66**
|
| MI+SM |
5.40±1.18Δ
|
After ligation of the left anterior descending artery for eight weeks, the number of PVBs obviously increased during the occlusion of the aortic arch for 5 s (**
P<0.01). The occurrence of PVBs significantly decreased with the use of SM, both in normal (*
P<0.05) and infarcted hearts (Δ
P<0.01). Data are expressed as mean±SD (n=15)
Fig.2
Monophasic action potentials (MAPs) and stretch-induced arrhythmias in chronically infarcted rat hearts
Upper figure: left ventricular pressure (LVP); Middle figure: MAP; Lower figure: ECG. (a) The occurrence of premature ventricular beats (PVBs) (thick arrows) during stretching in normal hearts; (b) Streptomycin (SM) inhibited the appearance of PVBs induced by stretching; (c) The number of PVBs increased significantly after ligation of the left anterior descending artery for eight weeks, and ventricular tachycardia (arrows with open hole) also occurred; (d) The occurrence of arrhythmias (arrows with dotted line) was reduced by pretreatment with SM
During the occlusion of the aortic arch for 5 s, the number of VT in the control group was 1, whereas it increased to 7 in the MI group. After treatment with SM, the number of VT decreased to 0 in the SM group and to 3 in the MI+SM group. The incidence of VT was higher than that in the control group (
P<0.05). SM decreased the occurrence of VT, but there was no significant difference between incidence in the MI+SM group and the MI group (
P>0.05) (Table
2).
Table 2
Effect of streptomycin (SM) on the occurrence of ventricular tachycardia (VT) during stretching for 5 s
| Group |
Number of hearts with VT |
Number of hearts without VT |
| Control |
1 |
14 |
| SM |
0 |
15 |
| MI |
7*
|
8 |
| MI+SM |
3 |
12 |
The number of hearts with VT increased with stretching after ligation of the left anterior descending artery for eight weeks (*
P<0.05). SM decreased the occurrence of VT, but the difference was not significant (P>0.05)
4. Discussion
The present study found that the application of SM
in vivo could inhibit changes in MAPD
90 and decrease the incidence of PVBs induced by stretching of chronically infarcted rat hearts. However, SM did not reduce the occurrence of VT, indicating the involvement of SACs in the genesis of stretch-induced PVBs in chronic MI, but suggesting a more complicated situation for stretch-induced VT.
4.1. Changes in MAPD90 and arrhythmias induced by stretching
Changes in MAPD caused by transiently increasing LV pressure have been reported in the normal myocardium of many species (Reiter et al.,
1997; Takagi et al.,
1999; Lerman et al.,
2001). Consistent with the results of earlier studies, the MAPD
90 of the LV was shortened during transient aortic occlusion in this study. Furthermore, we found that the stretch-induced decrease in MAPD
90
in vivo also had happened in the chronic MI rat hearts. By using a slice from the LV after MI treatment for five weeks, Kiseleva et al. (
2000) found that the sensitivity of the membrane potential and depolarization to stretch increased. Fu et al. (
2007) verified that the phenomenon of MEF increased in acute myocardium infarction (AMI) as well. The contraction force of ischaemic and infarcted myocardium was lower than that of surrounding normal myocardium after MI. So, stretch exists between the normal and ischemic myocardia. Likewise, stretch also exists between the infarcted and ischemic myocardia in chronic MI. In the present study, more PVBs and VT occurred in infarcted myocardium with occlusion of the aorta, which is consistent with previous results (Horner et al.,
1996).
4.2. Effects of the application of SM in vivo on MAPD90
In single cardiomyocytes (Belus and White,
2003) and multicellular preparations (Eckardt et al.,
2000; Spangenburg and McBride,
2006), SM has been verified to be an effective blocker of SACs. This study confirmed that the stretch-induced decrease in MAPD
90 in chronic MI could be pharmacologically inhibited by an SAC blocker. SM had no effect on the MAPD
90 under basic conditions, but effectively inhibited the shortening of the MAPD
90 induced by transient aortic occlusion in both normal and chronically infarcted hearts. Based on these results and the characteristics of MEF, we inferred that: (1) in chronic MI, the activities of SACs were involved in the stretch-induced changes in the MAPD
90; (2) treatment with SM
in vivo can inhibit the activities of SACs.
4.3. Effects of the application of SM in vivo on SIAs
SM showed different effects on PVBs and VT. SM decreased the occurrence of PVBs caused by stretching. The Ca
2+ cycle and the activities of SACs are the main mechanisms of SIAs (Lab,
1996). In short, the activities of SACs during stretching caused an increase in intracellular Ca
2+ ([Ca
2+]
i), which leads to after-depolarization and subsequent triggered activity (Fu et al.,
2007). The mechanism of the anti-arrhythmia effect of SM might be that by blocking SACs, it inhibits the increase in [Ca
2+]
i.
On the other hand, SM did not alter the occurrence of VT induced by aortic occlusion. There may have been several possible reasons for this: (1) the lack of a significant difference in this study may have been because of the small sample size; (2) absorption efficiency may have varied among different tissues following the application of SM
in vivo, causing our results to differ from those of previous studies (Yeung et al.,
2005; Spangenburg and McBride,
2006); (3) besides SACs, a more complicated mechanism may be involved in stretch-induced VT.
5. Conclusions
Stretch-induced MAPD
90 changes and arrhythmias via MEF were observed in chronically infarcted myocardium. The use of SM
in vivo decreased the incidence rate of PVBs but not VT, suggesting the involvement of SACs in MEF but that there might be other mechanisms involved in the formation of VT in chronically infarcted myocardium.
The results of our study revealed increased MEF in chronic MI and the effectiveness of SM
in vivo in minimizing certain electrophysiological changes during stretching. However, because of the risk of hearing and kidney damage caused by SM, further pharmacological studies are needed to find SM analogues which are safe for clinical application. Moreover, other studies will be necessary to explore further the mechanism of action of SIAs because other mechanosensitive ion channels in myocardium (Healy and McCulloch,
2005) may also be involved in stretch-induced electrical activity abnormalities, especially VT.
Acknowledgements
The authors thank Prof. Shu-xue LI (Heilongjiang University of Traditional Chinese Medicine, Harbin, China) for some reviews on technology.
* Project supported by the National Natural Science Foundation of China (No. 81301343)Compliance with ethics guidelines Jun-xian CAO, Lu FU, Qian-ping GAO, Rong-sheng XIE, and Fan QU declare that they have no conflict of interest.References
[1] Ashikaga, H., Mickelsen, S.R., Ennis, D.B., 2005. Electromechanical analysis of infarct border zone in chronic myocardial infarction.
Am J Physiol Heart Circ Physiol, 289(3):1099-1105.

[2] Belus, A., White, E., 2003. Streptomycin and intracellular calcium modulate the response of single guinea-pig ventricular myocytes to axial stretch.
J Physiol, 546((Pt 2)):501-509.

[3] Bertini, M., Ng, A.C., Borleffs, C.J., 2010. Longitudinal mechanics of the periinfarct zone and ventricular tachycardia inducibility in patients with chronic ischemic cardiomyopathy.
Am Heart J, 160(4):729-736.
[4] Cooper, P.J., Kohl, P., 2005. Species- and preparation-dependence of stretch effects on sino-atrial node pacemaking.
Ann N Y Acad Sci, 1047(1):324-335.
[5] Curtis, M.J., Hancox, J.C., Farkas, A., 2013. The Lambeth Conventions (II): guidelines for the study of animal and human ventricular and supraventricular arrhythmias.
Pharmacol Ther, 139(2):213-248.
[6] Eckardt, L., Kirchhof, P., Monnig, G., 2000. Modification of stretch-induced shortening of repolarization by streptomycin in the isolated rabbit heart.
J Cardiovasc Pharmacol, 36(6):711-721.
[7] Eckardt, L., Kirchhof, P., Breithardt, G., 2001. Load-induced changes in repolarization: evidence from experimental and clinical data.
Basic Res Cardiol, 96(4):369-380.
[8] Fu, L., Cao, J.X., Xie, R.S., 2007. The effect of streptomycin on stretch-induced electrophysiological changes of isolated acute myocardial infarcted hearts in rats.
Europace, 9(8):578-584.
[9] Garan, A.R., Maron, B.J., Wang, P.J., 2005. Role of streptomycin-sensitive stretch-activated channel in chest wall impact induced sudden death (commotio cordis).
J Cardiovasc Electrophysiol, 16(4):433-438.
[10] Haugaa, K.H., Smedsrud, M.K., Steen, T., 2010. Mechanical dispersion assessed by myocardial strain in patients after myocardial infarction for risk prediction of ventricular arrhythmia.
JACC Cardiovasc Imaging, 3(3):247-256.
[11] Healy, S.N., McCulloch, A.D., 2005. An ionic model of stretch-activated and stretchmodulated currents in rabbit ventricular myocytes.
Europace, 7((Suppl. 2)):128-134.
[12] Horner, S.M., Dick, D.J., Murphy, C.F., 1996. Cycle length dependence of the electrophysiological effects of increased load on the myocardium.
Circulation, 94(5):1131-1136.
[13] Kim, Y., White, E., Saint, D.A., 2012. Increased mechanically-induced ectopy in the hypertrophied heart.
Prog Biophys Mol Biol, 110((2-3)):331-339.
[14] Kiseleva, I., Kamkin, A., Wagner, K.D., 2000. Mechanoelectric feedback after left ventricular infarction in rats.
Cardiovas Res, 45(2):370-378.
[15] Lab, M.J., 1996. Mechanoelectric feedback (transduction) in heart: concepts and implications.
Cardiovasc Res, 32(1):3-14.
[16] Lab, M.J., 2006. Mechanosensitive-mediated interaction, integration, and cardiac control.
Ann N Y Acad Sci, 1080(1):282-300.
[17] Lerman, B.B., Engelstein, E.D., Burkhoff, D., 2001. Mechanoelectrical feedback: role of β-adrenergic receptor activation in mediating load-dependent shortening of ventricular action potential and refractoriness.
Circulation, 104(4):486-490.
[18] Ninio, D.M., Saint, D.A., 2008. The role of stretch-activated channels in atrial fibrillation and the impact of intracellular acidosis.
Prog Biophys Mol Biol, 97(2-3):401-416.
[19] Ravens, U., 2003. Mechano-electric feedback and arrhythmias.
Prog Biophys Mol Biol, 82((1-3)):255-266.
[20] Reiter, M.J., Landers, M., Zetelaki, Z., 1997. Electrophysiological effects of acute dilatation in the isolated rabbit heart: cycle length-dependent effects on ventricular refractoriness and conduction velocity.
Circulation, 96(11):4050-4056.
[21] Salmon, A.H., Mays, J.L., Dalton, G.R., 1997. Effect of streptomycin on wall-stress-induced arrhythmias in the working rat heart.
Cardiovasc Res, 34(3):493-503.
[22] Spangenburg, E.E., McBride, T.A., 2006. Inhibition of stretch-activated channels during eccentric muscle contraction attenuates p70
S6K activation.
J Appl Physiol, 100(1):129-135.
[23] Taggart, P., Lab, M.J., 2008. Cardiac mechano-electric feedback and electrical restitution in humans.
Prog Biophys Mol Biol, 97((2-3)):452-460.
[24] Takagi, S., miyazaki, T., Moritani, K., 1999. Gadolinium suppresses stretch-induced increases in the differences in epicardial and endocardial monophasic action potential durations and ventricular arrhythmias in dogs.
Jpn Circ J, 63(4):296-302.
[25] Thygesen, K., Uretsky, B.F., 2004. Acute ischaemia as a trigger of sudden cardiac death.
Eur Heart J Suppl, 6((Suppl. D)):D88-D90.
[26] Trapero, I., Chorro, F.J., Such-Miquel, L., 2008. Effect of streptomycin on stretch-induced change in myocardial activation during ventricular fibrillation.
Rev Esp Cardiol, 61(2):201-205.
[27] Wang, J.A., 2012. Progress and challenges in the cardiovascular field.
J Zhejiang Univ-Sci B (Biomed & Biotechnol), 13(8):587-588.
[28] Yeung, E.W., Whitehead, N.P., Suchyna, T.M., 2005. Effects of stretch-activated channel blockers on [Ca
2+]
i and muscle damage in the mdx mouse.
J Physiol, 562(Pt 2):367-380.

Open peer comments: Debate/Discuss/Question/Opinion
<1>