1. Introduction
Ubiquitin (Ub) is a highly conserved 76-residue protein, which can be covalently joined to the lysine (K) residues of target proteins as a single Ub or poly-Ub chain in eukaryotes (Jentsch,
1992; Grisvard et al.,
2010). Protein ubiquitination is an essential post-translational modification system that serves as a signaling mechanism in diverse cellular events, including ribosome biogenesis (Finley et al.,
1989), DNA repair (Jentsch et al.,
1987; Hofmann and Pickart,
1999; Pastushok and Xiao,
2004), protein endocytosis (Galan and Haguenauer-Tsapis,
1997), cell-cycle control (Wei et al.,
2004), nuclear factor kappa B (NF-κB)-dependent signal transduction (Chen et al.,
1996; Tokunaga et al.,
2009), and other cellular processes (Finley et al.,
1987). Ubiquitination is accomplished by a series of three enzymatic systems including Ub-activating enzyme (E1), Ub-conjugating enzyme (Ubc or E2), and Ub ligase (E3). In addition, Ub molecules can be linked to each other through an isopeptide bond formed between the C-terminal Gly76 of a donor and one of seven lysine residues of an acceptor (McKenna et al.,
2003), leading to the formation of poly-Ub chains (Chau et al.,
1989). Conventional poly-Ub via Gly76-Lys48 is the principal signal for proteolysis through 26S proteasomes (Hochstrasser,
1996), while Gly76-Lys63 poly-Ub regulates diverse activities in a non-proteolytic way (Pickart,
2001). So far, Ubc13 is the only known Ubc capable of catalyzing the Lys63-linked polyubiquitylation reaction and this function requires interaction with the Ubc variant (Uev, which shares a similar sequence and structure with the E2 Ub-conjugating enzymes, but lacks the active cysteine residue); thus, it is unlike other Ubc proteins that catalyze a conventional Lys48-linked polyubiquitylation reaction (Hofmann and Pickart,
1999; McKenna et al.,
2001).
Ubc13 was originally identified as
bendless because its loss-of-function mutation affected neuronal synaptic connectivity in
Drosophila melanogaster (Muralidhar and Thomas,
1993; Oh et al.,
1994). The yeast
ubc13 null mutant displays sensitivity to DNA-damaging agents such as ultraviolet (UV) or methylmethane sulfate (MMS) and a high spontaneous mutation rate (Brusky et al.,
2000). Lys63-linked chains catalyzed by the Ubc13-Uev (Mms2) complex are required for error-free DNA damage tolerance (DDT, also known as post-replication repair (PRR)) by polyubiquitinating the proliferating cell nuclear antigen (PCNA) (Broomfield et al.,
1998;
2001; Hofmann and Pickart,
1999; Hoege et al.,
2002) in
Saccharomyces cerevisiae. The human Ubc13 protein (hUbc13) has also been cloned (Yamaguchi et al.,
1996). The transcription factor NF-κB, which is sequestered in the cytoplasm via interacting with an inhibitory protein of the IκB family, controls many processes including immunity, inflammation, and apoptosis. IκB proteins can be rapidly phosphorylated by an IκB kinase (IKK) complex and then degraded (Weil and Israel,
2004; Chen,
2005). hUbc13-Uev1A complex is involved in activating the tumor necrosis factor receptor-associated factor 6 (TRAF6)-mediated IKK by catalyzing the synthesis of Lys63-linked poly-Ub chains, which perform polyubiquitination of TRAF6 (Chen,
2012). Mouse Ubc13 is able to functionally complement the budding yeast
ubc13 null mutation (Ashley et al.,
2002). Ubc13 appears to be essential for the thymocyte T cell receptor (TCR)-mediated NF-κB activation at the early time points and transforming growth factor-β-activated kinase 1 (TAK1) phosphorylation in innate immune cells of mice (Sato et al.,
2005; Yamamoto et al.,
2006b). The deletion of the Ubc13 in mice results in severe loss of blood cells together with atrophy of the thymus and bone marrow, showing that Ubc13 also has a pivotal role in regulating hematopoiesis (Wu et al.,
2009). In both zebrafish and
Arabidopsis thaliana, there are two
UBC13 genes,
ubc13a and
ubc13b or
AtUBC13A and
AtUBC13B. All these four Ubc13 proteins can physically interact with human or yeast Mms2 and are able to functionally complement the yeast
ubc13 null mutant for spontaneous mutagenesis and sensitivity to DNA-damaging agents (Wen et al.,
2006; Li et al.,
2010); this implies the existence of a Lys63-linked polyubiquitylation reaction and error-free DDT pathway in zebrafish and
A. thaliana. ShUbc13 is also capable of physically interacting with ShUev and is probably involved in DNA damage response in the ciliate
Sterkiella histriomuscorum (Grisvard et al.,
2010). Recently, studies of the
Ubc13 gene in rice have been carried out.
OsUbc13 showed ubiquitous expression at a high level even under biotic or abiotic stresses, and its expression can complement the error-free PRR defects of the yeast
ubc13 null mutant. Moreover, OsUbc13 physically interacts with both Mms2 and Uev1A, and catalyzes K63-linked polyubiquitination
in vitro (Zang et al.,
2012).
Rice is a prominent model for monocotyledonous plants and one of the most important food crops (Yang et al.,
2011). However, only limited information is available on Ubc13 in plant species. In this study, we cloned and analyzed the molecular characterization of
OsUbc13, and studied its subcellular localization and interacting proteins, providing a foundation for further investigating its function and molecular mechanism.
2. Materials and methods
2.1. Plant materials and treatments
The mature seeds of a rice variety, Nipponbare, were dehulled and sterilized in 10% (0.1 g/ml) NaClO for 15 min, germinated and grown in semi-solid 1/2 MS medium (Murashige and Skoog,
1962) at 28 °C with a photoperiod of 16 h light/8 h dark for two weeks. Seedlings were transferred to Yoshida solution (Yoshida et al.,
1976) for two weeks, and finally transferred outside and grown naturally in a field. Plant components at different developmental stages (roots and leaves from the four-week-old seedlings; stem, stamen, pistil, lemma, and palea from the panicles at 15 d after flowering) were collected for real-time polymerase chain reaction (PCR).
To analyze the expression in response to hormone and abiotic stresses, an embryonic cell culture line from the japonica line Nipponbare under culture conditions of 28 °C in the darkroom was used. For low-temperature treatment, the embryonic cell cultures were directly transferred to a 4 °C incubator from the normal growth conditions. The culture at 28 °C in the darkroom was employed as the control. For mannitol, H
2O
2, MMS, abscisic acid (ABA), and NaCl treatments, the embryonic cell cultures were transferred from semi-solid N6 medium to the liquid N6 medium supplemented with 300 mmol/L mannitol, 20 mmol/L H
2O
2, 0.01% MMS, 100 μmol/L ABA, or 200 mmol/L NaCl, respectively. The embryonic cell cultures grown in the same liquid N6 medium without supplementary components were used as controls. All the cultures, except for the control at 0 h, were placed in a shaker at 200 r/min at 28 °C and sampled at 2, 4, 8, 12, and 24 h. Each treatment was repeated three times.
All materials were quickly frozen in liquid nitrogen and then stored at −80 °C for RNA extraction.
2.2. Identification and cloning of OsUbc13 gene
OsUbc13, corresponding to protein spot 5002, was identified from the specific-organ-inducing medium-induced somatic root and shoot regeneration system in our previous study (data not shown). Analysis of resultant peptides obtained from mass spectrum assay was performed by Expasy (
http://prosite.expasy.org/;
http://web.expasy.org/compute_pi/).
The full length complementary DNA (cDNA) of
OsUbc13 gene was amplified by reverse transcription PCR (RT-PCR). The gene-specific primers for RT-PCR were Ubc13-F (5'-
GAATTCATGGCCAACAGCAACCTCC-3') and Ubc13-R (5'-
CCCGGGTTATGCACCGCTGGCATACA-3') with
EcoRI and
SmaI restriction sites, respectively. Purified PCR products were cloned into pMD18-T vector (TaKaRa) and then confirmed by sequencing (Invitrogen). The resulting plasmid was called pMDUbc13.
2.3. Phylogenic analysis of Ubc13 proteins
For analysis of amino acid sequence identity, DNAMAN software was used. Amino acid sequences of Ubc13 from different species were aligned using ClustalW2. Analysis of phylogeny was performed using MEGA4 via the neighbor-joining method (Tamura et al.,
2007). All the amino acid sequences (given in Online Resource 1) were retrieved from the NCBI database (
http://www.ncbi.nlm.nih.gov/).
2.4. Expression analysis of OsUbc13 gene by real-time PCR
Total RNA was isolated from rice materials using Trizol Reagent (Invitrogen, USA) according to the manufacturer’s protocol. The first-strand cDNAs were synthesized using the Moloney murine leukemia virus (M-MLV) first-strand synthesis system with 1 μg of freshly extracted RNA (Promega). The SYBR Premix Ex Taq Kit (TaKaRa) was used for real-time PCR analysis, with the primers qUbc13-F (5'-ATGGCCAACAGCAACCTCC-3') and qUbc13-R (5'-TTATGCACCGCTGGCATACA-3'), qActin-F (5'-GACTCTGGTGATGGTGTCAGC-3') and qActin-R (5'-GGCTGGAAGAGGACCTCAGG-3'). The PCR program and the calculation method of relative expression levels of
OsUbc13 in different tissues or under different conditions were as previously described (Wang Y. et al.,
2012), with the rice
Actin I gene as a reference. Here, ∆∆
C
T=(
C
T(Ubc13, test)−
C
T(Actin I, test))−(
C
T(Ubc13, calibrator)−
C
T(Actin I, calibrator)), where the
C
T (cycle threshold) values for both target and reference genes are the means of the triplicate independent PCRs. Here, the roots or embryonic cell cultures without treatment at 0 h were set as calibrators.
2.5. Subcellular localization of OsUbc13
The subcellular localization of OsUbc13 was analyzed by constructing a fusion protein, in which the
OsUbc13 coding sequence (CDS) was fused to green fluorescent protein (GFP). The
OsUbc13 CDS was amplified from pMDUbc13 with the primers slUbc13-F (5'-ATGGCCAACAGCAACCTC-3') and slUbc13-R (5'-TTATGCACCGCTGGCATAC-3') and then inserted into the pGWC, which had been digested by Eam1105I. The fragment was then cloned into the pMDC43 vector via LR reaction (Gateway Technology, Catalog Nos. 12535-019 and 12535-027) to be fused to the downstream of GFP, resulting in pMDC43-Ubc13, denominated as pGFP:Ubc13 (given in Online Resource 2). This final construct was transformed in cultured tobacco BY-2 protoplasts using a procedure as reported previously (Sheen,
2001; Lee et al.,
2008; Silva et al.,
2010) with some modifications. Briefly, the suspended BY-2 protoplasts were subcultured after every 3–4 d by transferring 10 ml cells into 40 ml of fresh liquid MS medium. These suspended cells were then transferred to 50 ml conical tubes in a laminar flow hood and were centrifuged at about 500×
g for 5 min at 25 °C. After removing the supernatant, the sediment was dissolved in 25 ml of enzyme solution (1% (0.01 g/ml) cellulase, 0.1% (1 g/L) pectinase Y23, and 0.2% (2 g/L) driselase in 4.3 g/L MS salts, 0.4 mol/L sucrose, 0.5 g/L 2-morpholinoethanesulfonic acid (MES) hydrate, 0.25 g/L NH
4NO
3, 0.75 g/L CaCl
2·2H
2O, pH 5.7) and incubated in a shaker at 200 r/min at room temperature for 4–5 h. Then the mixture was centrifuged using a swinging bucket rotor at 80×
g for 15 min. The floating protoplast layer was transferred to a new tube and centrifuged for 5 min at 100×
g to collect BY-2 protoplasts. Finally, the transformed protoplasts were examined by confocal laser scanning microscopy (Zeiss LSM700).
2.6. Interaction assay of OsUbc13 by yeast two-hybrid
Matchmaker Two-Hybrid System 3 (Clontech,
http://www.clontech.com/), in which a bait gene is fused to the GAL4 DNA-binding domain while cDNA is fused to the GAL4 activation domain-AD (Fields and Song,
1989; Chien et al.,
1991), was used for protein interaction analysis. Briefly, the fragment that was obtained from pMDBTF3 was digested with
EcoRI and
SmaI was inserted into the pGBKT7 vector to generate construct pGUbc13 (given in Online Resource 2). An AD fusion library of rice calli in pGADT7 (pGADT7-library) was constructed in Clontech. The following test and re-test of positive two-hybrid interaction were performed as previously described (Wang Y. et al.,
2012).
3. Results
3.1. Sequence analysis of OsUbc13 protein
We identified a protein OsUbc13 corresponding to protein spot 5002 in 2D gels to be associated with somatic root and shoot regeneration in our previous study (data not shown). The complete coding sequence of
OsUbc13 contains eight exons and seven introns, and encodes a UBC homolog of 153 amino acids. As Zang et al. (
2012) proposed,
OsUbc13 (Os01g0673600) is the only
Ubc13 gene in rice because only this gene with a high degree of sequence identity was found using
Arabidopsis Ubc13 genes to BLAST the rice genomic database on NCBI. OsUbc13 shares high similarities with other homologs, ranging from 66.23% to 98.04%. Comparison of amino acid sequence revealed that all Ubc13s harbor the active site Cys-88 within the highly conserved UBC domain (Fig.
1a), indicating that UBC13 orthologs are derived from the same origin evolutionarily. Phylogenetic analysis (Fig.
1b) indicated that OsUbc13 is the most closely related to AtUBC13B, followed by AtUBC13A and animal Ubc13s, and distant from yeast ScUbc13. This finding is consistent with the previous report declaring that all except two
OsUbc13 introns are different from that of
AtUBC13B in the gene structure while there are obvious differences between
OsUbc13 and other
Ubc13s (Zang et al.,
2012).
Fig.1
Amino acid sequence alignment (a) and phylogenic analysis (b) of Ubc13s from different species
The gray and single overline in (a) indicates the conserved UBC domain of Ubc13s; the dark and single overline in (a) indicates the conserved cysteine residue of Ubc13s. Bootstrap values are shown at each node in (b). AtUBC13A: AEE36165; AtUBC13B: AEE29516.1; OsUbc13: NP_001043834.1; MmUbc13: AAK74128.1; HsUbc13: BAA11675.1; DrUbc13a: AAI64904.1; DrUbc13b: AAH53141.1; CeUbc13: NP_500272.2; DmUbc13a: AAF48338.1; DmUbc13b: ACZ95287.1; ScUbc13: CAA67806.1
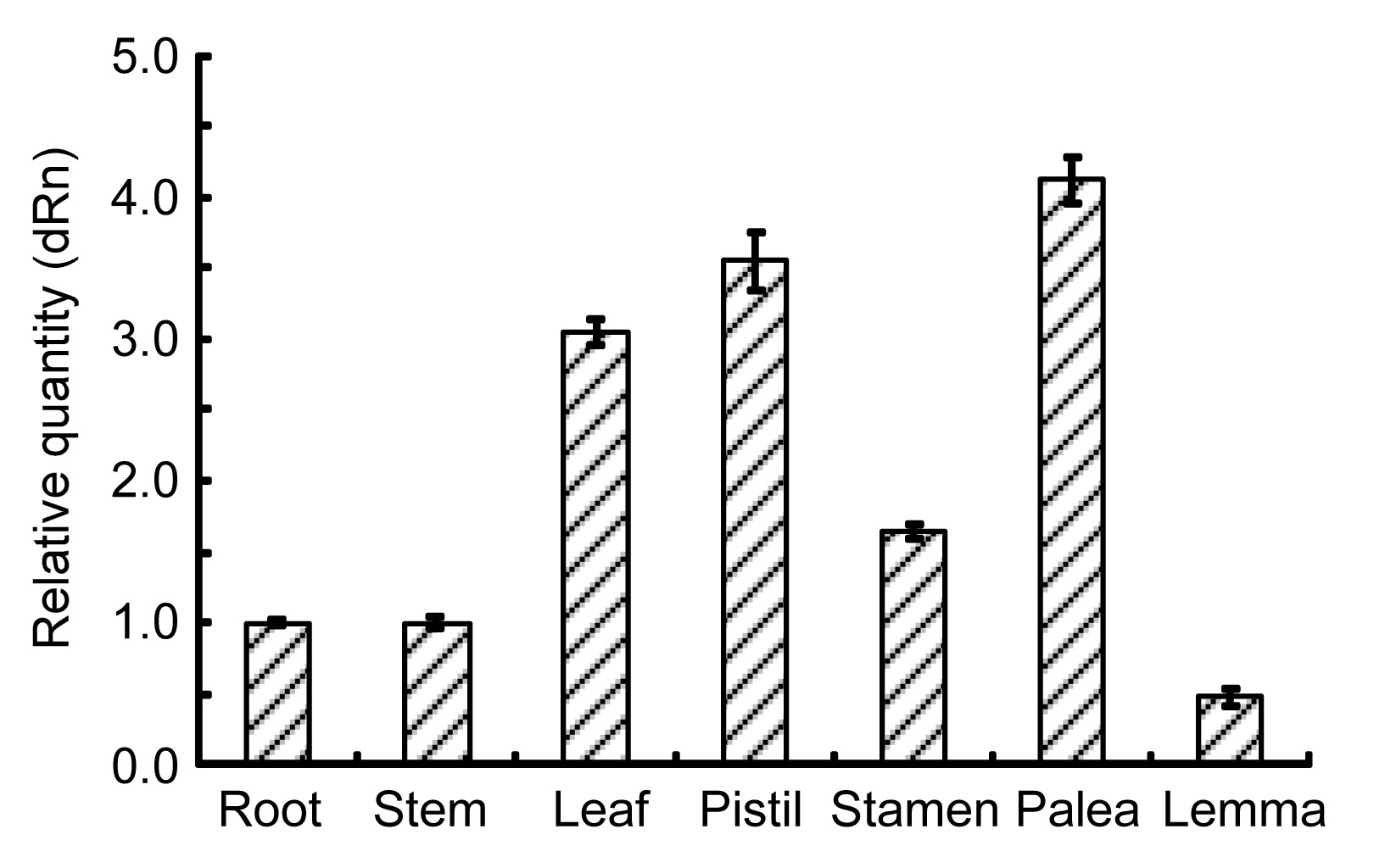
3.2. Expression of OsUbc13 in various rice tissues
To investigate the expression pattern of
OsUbc13, the relative abundance of mRNA in different tissues including root, stem, leaf, stamen, pistil, lemma, and palea was quantified. The results revealed that the endogenous
OsUbc13 was constitutively expressed in all tissues examined and the abundance level was highest in palea, followed by pistil, leaf, stamen, root, and stem, and lowest in lemma (Fig.
2). These results implied that the
OsUbc13 is important for both vegetative growth and reproductive development in rice, especially that of palea, pistil, and leaf.
Fig.2
Expression analysis of OsUbc13 in various rice tissues
The expression of OsUbc13 in root was set as 1. Data were expressed as mean±standard deviation (SD) of triplicate measurements
3.3. Expression of OsUbc13 in different stress conditions
In plants, many gene transcripts are very unstable, which is important for the tight control of their temporal and spatial expression patterns (Lee et al.,
2006). In order to determine the expression of
OsUbc13 in response to environmental conditions, its transcript level was analyzed by real-time PCR after treatment with low temperature (4 °C), MMS (0.01%), H
2O
2 (20 mmol/L), osmotic pressure (300 mmol/L mannitol), ABA (100 μmol/L), and NaCl (200 mmol/L). The results showed that the expression of
OsUbc13 was induced by low temperature (4 °C), 0.01% MMS, and 20 mmol/L H
2O
2 (Figs.
3a–3c), but repressed by 300 mmol/L mannitol, 100 μmol/L ABA, and 200 mmol/L NaCl (Figs.
3d–3f). For low temperature treatment, around four-fold transcripts in the treated samples were observed, compared with the controls at 2 h, the difference reached its maximum value at 12 h, but the level of expression decreased although it was still higher than that of the control at 24 h (Fig.
3a). For MMS and H
2O
2 treatments, the maximum level of expression was at 4 and 8 h, respectively. Moreover, the difference between treated samples and control was smaller in the case of MMS and H
2O
2 treatments (Figs.
3b–3c) than in low-temperature treatment (Fig.
3a). For the mannitol treatment, the transcripts were less in the samples treated than in the controls at all time points except at 2 h (Fig.
3d). The expression patterns of
OsUbc13 in response to ABA and NaCl were quite similar; however, the time point at which the maximum expression level occurred was different under different treatments. The former (under ABA treatment) was at 2 h and the latter (under NaCl treatment) was at 4 h (Figs.
3e–3f). Interestingly, the transcripts in the control cultures in liquid N6 medium (Figs.
3b–3f) fluctuated noticeably, compared with those cultured in solid N6 medium (Fig.
3a), indicating that culture conditions may affect the expression level of
OsUbc13.
Fig.3
Expression pattern of OsUbc13 in response to low temperature of 4 °C (a), 0.01% MMS (b), 20 mmol/L H2O2 (c), 300 mmol/L mannitol (d), 100 μmol/L ABA (e), and 200 mmol/L NaCl (f)
The detection was performed three times and the data were presented as mean±standard deviation (SD). The expression of OsUbc13 of control at 0 h was set as 1
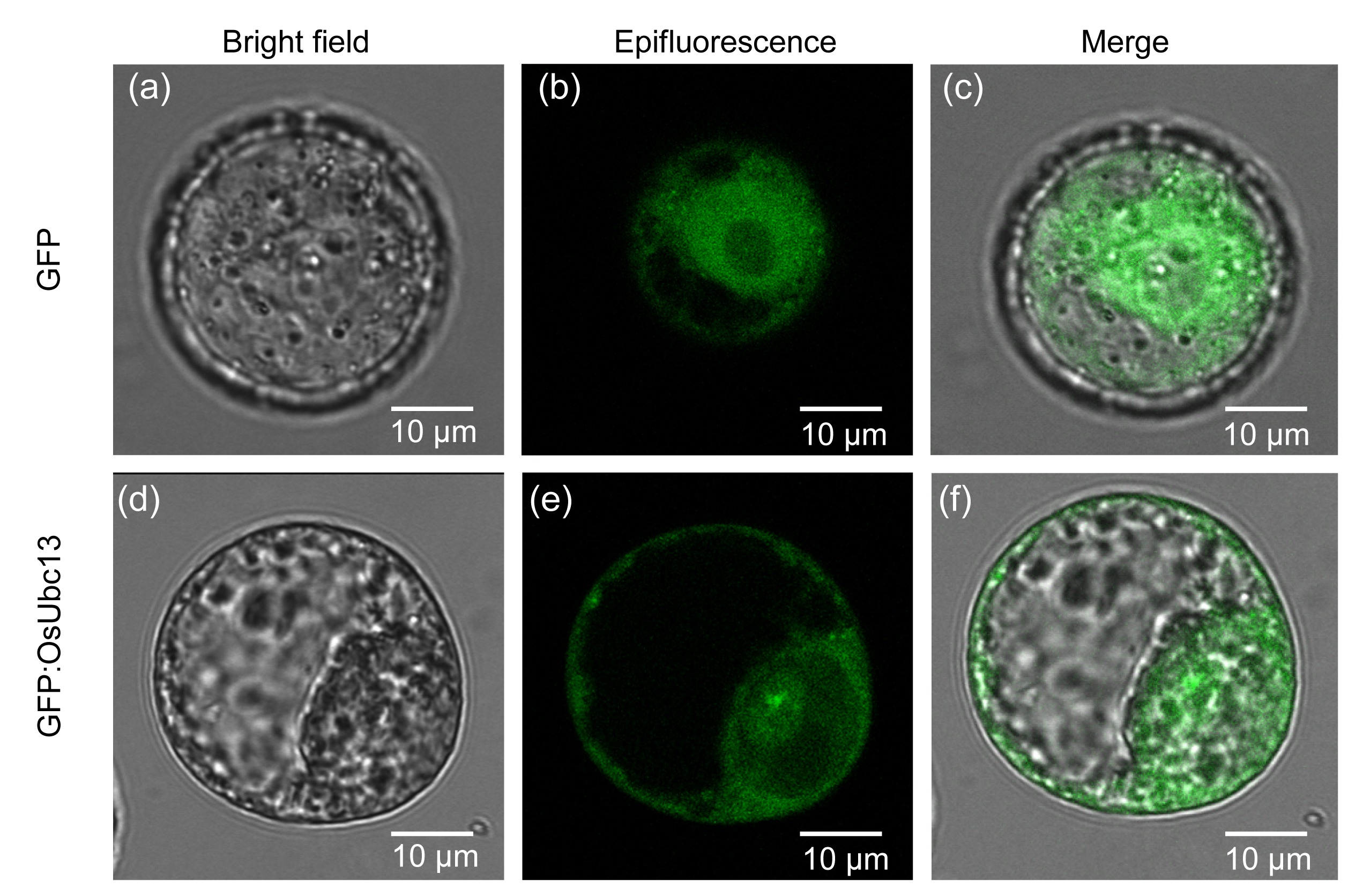
3.4. Localization of OsUbc13 in the plasma and nuclear membranes
To determine the subcellular localization of OsUbc13, the fusion construct pMDC43-Ubc13 containing GFP:OsUbc13 CDS and driven by 35S promoter was introduced into the protoplasts of cultured
Nicotiana benthamiana via polyethylene glycol (PEG)-mediated transformation. The non-fusion construct pMDC43 was used as a control. Here, the tobacco cell system as a model system was selected to study subcellular localization of OsUbc13 for its potential advantages over other cell cultures such as highly synchronized growth rate, high transformation efficiency, and easy monitoring and analysis (Geelen and Inzé,
2001; Tripathi et al.,
2013). The data revealed that OsUbc13 was probably localized in the plasma and nuclear membranes since the fluorescence signal of GFP:OsUbc13 was relatively intense in these areas (Figs.
4d–4f), while control GFP was localized in both the cytoplasm and nucleus (Figs.
4a–4c). The subcellular localization of OsUbc13 implied that OsUbc13 may function in the membrane system.
Fig.4
Subcellular localization of control GFP (a–c) and GFP:OsUbc13 (d–f) fusion proteins transiently expressed in the cultured BY-2 protoplast
GFP:OsUbc13 was localized in the plasma and nuclear membranes. Images were taken in bright field (a, d) for the cell outline and in dark field for green fluorescence (b, e). Images (c) and (f) are superimposed images between (a) and (b) as well as (d) and (e), respectively (Note: for interpretation of the references to color in this figure legend, the reader is referred to the web version of this article)
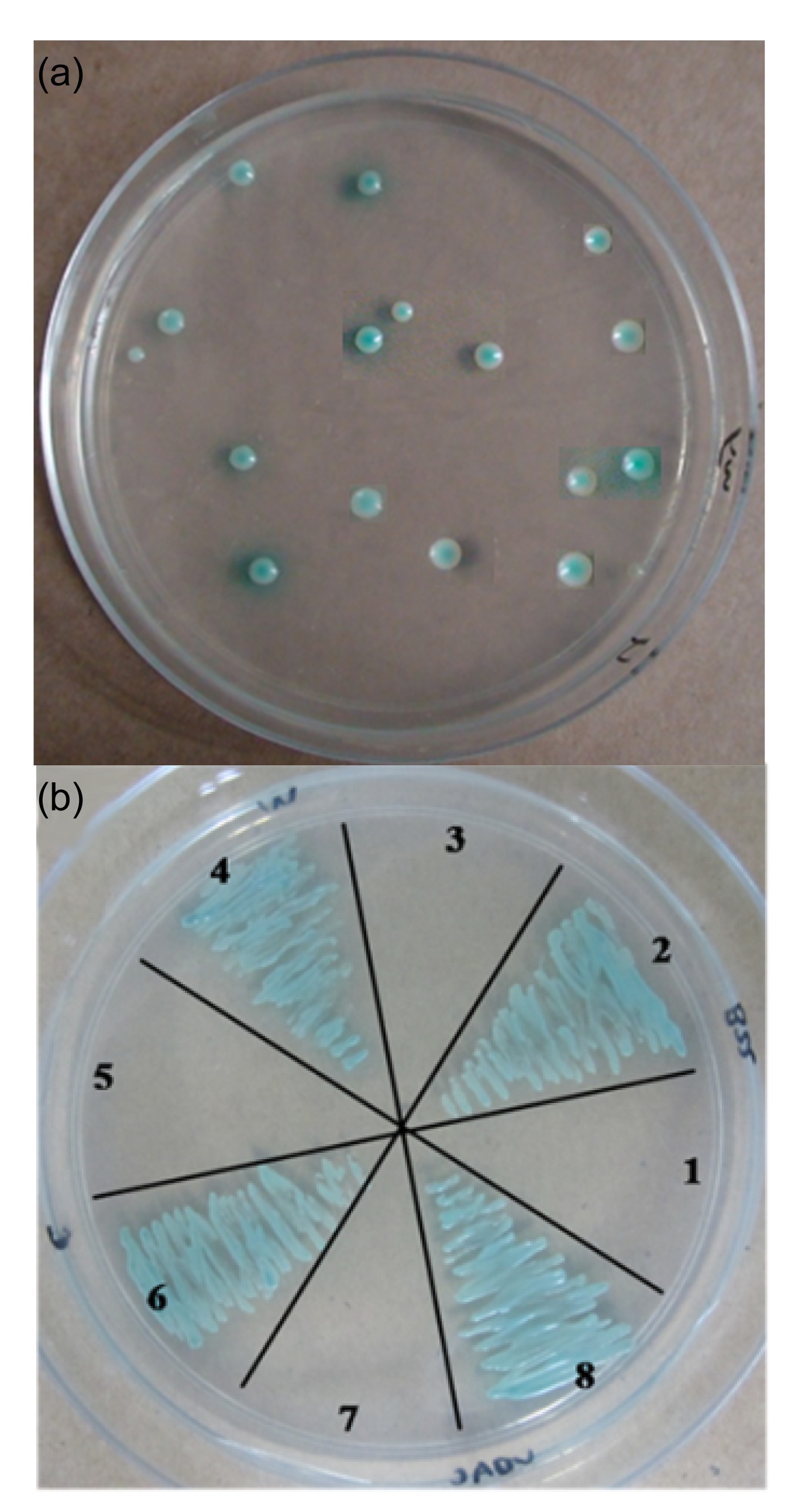
3.5. Potential OsUbc13-interacting partners in yeast two-hybrid assay
To provide insights into biological function and regulatory mechanism, yeast two-hybrid was performed to determine potential candidates. As the results showed, 81 colonies were positive (Fig.
5a). Out of them, 20 proteins (Table
1), which are responsible for the positive two-hybrid interaction, were identified via PCR colony-screening and DNA sequencing. The protein-protein interactions were verified by co-transformation in yeast AH109 with four genes (out of 20) as representatives. The four genes are
OsVDAC (Godbole et al.,
2003),
OsCROC-1 (Hofmann and Pickart,
1999),
OsB22EL8 (Yuan J. et al.,
2008), and
OsMADS1 (Prasad et al.,
2005) of which the function had been defined (Fig.
5b).
Fig.5
Yeast two-hybrid analysis with OsUbc13 as the bait
(a) The positive colonies on SD/-Ade/-His/-Leu/-Trp/X-α-gal plate with OsUbc13 as the bait. (b) The interactions were verified by co-transformation represented by OsB22EL8 (2), OsMADS1 (4), OsCROC-1 (6), and OsVDAC (8). 1, 3, 5, and 7 represent four negative controls of 2, 4, 6, and 8, respectively
Table 1
List of proteins interacted with OsUbc13 from yeast two-hybrid
| Function |
Protein description |
GenBank accession No. |
| Apoptosis |
Outer mitochondrial membrane protein porin (OsVDAC) |
Os09g0361400 |
| Hypersensitive response (HR)-like lesion-inducer family protein |
Os10g0445500 |
| Similar to cysteine protease inhibitor |
Os05g0409300 |
| DNA damage repair |
Polyubiquitin |
Os06g0681400 |
| Similar to CROC-1-like protein (OsCROC-1) |
Os12g0605400 |
| Similar to polyubiquitin |
Os04g0628100 |
| Similar to B22EL8 protein (OsB22EL8) |
Os05g0111300 |
| RNA polymerase subunit, RPB5/RPB6-like domain containing protein |
Os06g0618600 |
| Similar to CROC-1-like protein |
Os04g0684800 |
| Flower development |
Box protein (MADS box protein MADS1) (OsMADS1) |
Os03g0215400 |
| Arf GTPase activating protein family protein |
Os01g0951100 |
|
Oryza sativa SKP1 (S-phase kinase-associated protein 1)-like protein 1 (OSK1) |
Os05g0530500 |
| Immunological responses |
Pollen allergen Lol p2 family protein |
Os04g0317500 |
| Pollen allergen Lol p2 family protein |
Os06g0655200 |
| Catabolism |
Similar to 3-methylcrotonyl CoA carboxylase biotin-containing subunit |
Os12g0605800 |
| Splicing and cell cycle regulation |
Similar to U5 small nuclear ribonucleoprotein 200 kDa helicase |
Os02g0107000 |
| Other |
Similar to coatomer delta subunit (δ-coat protein (δ-COP)) |
Os08g0368000 |
| Similar to YyaF/YCHF TRANSFAC/OBG family small GTpase plus RNA binding domain TGS |
Os08g0199300 |
| Protein of unknown function DUF1685 family protein |
Os11g0282300 |
| Peptidase S14, ClpP family protein |
Os03g0411500 |
Another eight potential OsUbc13-interacting proteins (Table
1), along with these four confirmed genes, are associated with apoptosis (Yang et al.,
1997; Kim et al.,
2009; Hashimoto et al.,
2012; Wang W.M. et al.,
2012), DNA repair and damage tolerance (Parusel et al.,
2006), and floral organ development (Dai et al.,
2002; Takacs et al.,
2012). The remaining eight proteins (Table
1) are involved in immunological responses (Petersen et al.,
2006; Schweimer et al.,
2008), catabolism (Song et al.,
1994; Weaver et al.,
1995; Guan et al.,
1999; Obata et al.,
2001), splicing and cell-cycle regulation (Lauber et al.,
1996; Ehsani et al.,
2013). These results indicated that OsUbc13 might function in diverse biological processes.
4. Discussion
UBC13 is a Ub-conjugating enzyme involved in error-free DDT (or PRR) by catalyzing the synthesis of Lys63-linked poly-Ub chains together with a Uev (Broomfield et al.,
1998) and a variety of stresses (Hiraishi et al.,
2006). In this study, we cloned and characterized
OsUbc13, which encodes a Ub-conjugating protein. Although the amino acid sequence comparison of rice Ubc13 with Ubc13s from other species (Fig.
1a) is similar to previous results (Zang et al.,
2012), the range of UBC13 orthologs is wider and all Ubc13s harbor the active site Cys-88 within the highly conserved UBC domain; this indicates that UBC13 orthologs from entire eukaryotic organisms are derived from the same origin evolutionarily.
OsUbc13 was constitutively expressed in all the tissues and organs examined (Fig.
2), implying that OsUbc13 may exert important roles in the growth and development of rice. It is worth considering that both
AtUBC13s and
OsUbc13 were not only expressed ubiquitously in different tissues but also kept at a relatively stable transcript level in the previous report (Wen et al.,
2006; Zang et al.,
2012), and this is a little different from our results. Our data showed that
OsUbc13 was expressed abundantly in palea, pistil, leaf, and stamen, compared with its lower expression level in root, stem, and lemma (Fig.
2). In our opinion, the differences, if any, are at least partly due to different research methods (e.g., the detection technology or the method of data analysis). A cross-database search on the Genevestigator (
https://www.genevestigator.com/gv/doc/plant/introduction.jsp) was performed to investigate the expression profiles of
OsUbc13 in the previous report while our results were obtained via real-time PCR using three independent sets of biological samples.
Previous reports showed that the disruption of the
Ubc13 gene results in an increased killing by MMS and UV irradiation as well as an increased spontaneous mutation rate in yeast (Brusky et al.,
2000), implying that the expression of
Ubc13 is susceptible to adverse environments, especially those causing damage to DNA. Hence, in order to identify the response of
OsUbc13 expression to different stresses, real-time PCR was performed to check the expression pattern of
OsUbc13 under conditions of low temperature 4 °C, mannitol, H
2O
2, MMS, ABA, and NaCl. It is reasonable to assume that the expression of
Ubc13 may be regulated by stresses since Ubc13 is mainly involved in stress responses such as DNA damage. In fact, yeast and zebrafish
Ubc13s are induced by different DNA damaging agents such as MMS (Brusky et al.,
2000; Li et al.,
2010). Similarly, MMS up-regulated the expression of
OsUbc13 (Fig.
3b), and the expression levels under treatments of low temperature and H
2O
2 were also higher than those under normal conditions (Figs.
3a and 3c). In contrast, its expression was down-regulated by ABA and NaCl (Figs.
3e and 3f). For osmotic pressure treatment, the expression level was increased at 2 h and after that it decreased continually, compared with the control (Fig.
3d); this was slightly different from the expression pattern of
OsUbc13 to other stress treatments. However, the transcript level of
OsUbc13 appears to fluctuate under different stresses in this study, and this is in contrast to previous results from
Arabidopsis and rice (Wen et al.,
2006; Zang et al.,
2012). The differences are probably attributable to the experimental methods. Here, we used embryonic cell culture for the analysis of
OsUbc13 gene expression in response to various stresses, and detected its expression at different time points. In conclusion, our data showed that
OsUbc13 has an important role in plant responses to stress tolerance and can respond actively to negative circumstances by changing its transcripts. It should be noted that even the “untreated” controls, all of which were cultured in liquid N6 medium, are rather different (Figs.
3b–3f). We think the reason for the differences is probably the different culture conditions. The cultures that had not been placed in the shaker were sampled as controls at 0 h, while controls at the other time points were cultured in the shaker at 200 r/min, implying that culture conditions may affect the gene expression level.
It is important to determine the subcellular localization of corresponding proteins to understand the function of a gene as a whole. In yeast (Ulrich and Jentsch,
2000), mammals (Andersen et al.,
2005), and zebrafish (Li et al.,
2010), Ubc13 was considered to be a cytoplasmic protein. In the single-celled organism, the ciliate
S. histriomuscorum, Ubc13 protein shows three main locations: nucleus, cilia, and plasma membrane (Grisvard et al.,
2010). However, there has been no report on cellular localization of a plant Ubc13 protein. In this study, we are the first to demonstrate that the fluorescence of GFP:OsUbc13 was more strongly close to the plasma and nuclear membranes (Fig.
4e), implying that OsUbc13 may function in the membrane system; this is a difference from the previous results from yeast, animals, and
S. histriomuscorum (Ulrich and Jentsch,
2000; Andersen et al.,
2005; Grisvard et al.,
2010; Li et al.,
2010). For further studies, it will be essential to confirm the localization of OsUbc13 via adding a membrane-specific marker.
To understand further the functions of Ubc13-mediated protein ubiquitination, it is important to study Ubc13-interacting proteins. About 20 proteins (Table
1), most of which have not been described previously, are identified as potential OsUbc13-interacting partners. We selected four genes to confirm the interactions (Fig.
5b) since these genes are representative in the several types of genes listed in Table
1 and their functions have already been studied systematically. For instance, Ub-conjugating E2 enzyme variant (Uev) CROC-1, which was originally identified on the basis of its capacity to activate transcription of c-Fos (Rothofsky and Lin,
1997), lacks a critical cysteine residue essential for the catalytic activity of E2 enzymes. Uev can functionally complement the yeast
mms2 null mutant from killing by DNA-damaging agents, including MMS and UV (Xiao et al.,
1998). In
A. thaliana, all four Uev1 proteins can interact with AtUBC13 or Ubc13 from yeast or human to form a stable complex. Furthermore, when treated with a DNA-damaging agent, both seeds and pollen of
uev1d plants germinated poorly (Wen et al.,
2008). Zang et al. (
2012) reported that OsUbc13 physically interacts with Mms2 and Uev1A, which are hUbc13 partners, and catalyzes K63 polyubiquitination
in vitro. We demonstrated here that OsUbc13 physically interacted with OsCROC-1 (Fig.
5b; Table
1), which is essential for Ubc13-mediated Lys63 polyubiquitylation (Hofmann and Pickart,
1999; McKenna et al.,
2001; Wen et al.,
2006); this once again implies the existence of error-free DDT in plants and the critical role of OsUbc13 in tolerance to DNA damage during seed and pollen germination.
Apoptosis plays an important role during development, homeostasis, and in many diseases such as cancer (Steller,
1995). OsVDAC, which is known to be involved in apoptosis, is another confirmed OsUbc13-interacting protein here. VDAC is a mitochondrial outer membrane ion channel and a critical participant in the apoptosis of animal and plant cells (Al Bitar et al.,
2003; Godbole et al.,
2003). In plants, over-expression of rice VDAC (
OsVDAC4) induces apoptosis (Godbole et al.,
2011). In animal cells, knocking down
VDAC1 by small interfering RNA attenuates endostatin (ES)-induced apoptosis, while over-expression of
VDAC1 enhances the sensitivity of endothelial cells to ES (Yuan S. et al.,
2008). The previous study indicated that apoptosis may be induced via transmembrane receptors that belong to the tumor necrosis factor (TNF) receptor family (Susin et al.,
1997) and TNF receptor signaling has been closely linked to TAK1, which needs the involvement of Ubc13 in phosphorylation (Yamamoto et al.,
2006a). The yeast two-hybrid results showed that OsUbc13 interacted with OsVDAC (Fig.
5b; Table
1), indicating that OsUbc13 might function in apoptosis via TNF receptor signaling.
The third confirmed OsUbc13-interacting protein OsB22EL8 is a kind of metallothionein (MT) protein. MTs play important roles in maintaining homeostasis and detoxification of metals in both plants and animals; they are known as a kind of low molecular mass and cysteine-rich metal-binding protein (Yuan J. et al.,
2008). In plants, studies on the responses of MTs to stress conditions, such as drought and reactive oxygen species (ROS), have also been conducted widely (Akashi et al.,
2004; Wong et al.,
2004). It is well known that both abiotic and biotic stresses can induce the increased ROS such as superoxide, hydrogen peroxide, singlet oxygen, and hydroxyl radicals at certain stages of stress exposure. These ROS may cause non-specific oxidation of proteins and membrane lipids or DNA injury (Pastori and Foyer,
2002; Apel and Hirt,
2004). MTs can improve the stress tolerance of plants by participating in a complex ROS scavenging system to minimize the effects of oxidative stress (Yang et al.,
2009). We speculate that OsUbc13 might participate in DNA protection by an ROS-scavenging pathway considering the fact that OsUbc13 interacted with OsB22EL8 (Fig.
5b; Table
1).
OsUbc13 is also probably involved in floral organ identity. OsMADS1, a rice MADS-box factor, which is the last OsUbc13-interacting protein identified in our study (Fig.
5b; Table
1), controls both differentiation and proliferation of specific lemma and palea cell types and is an early-acting regulator of inner floral organs (Prasad et al.,
2005). Both the gain-of-function and the loss-of-function of OsMADS1 can lead to severe defects in the specification of lemma and palea (Agrawal et al.,
2005). Previous studies on Ubc13 focused on error-free DDT pathways and other cellular processes, and our findings that OsUbc13 interacted with OsMADS1 could open a new research area of Ubc13 in plant flower development.
However, we selected only four of the candidate genes to confirm the interactions without re-testing the others such as hypersensitive response (HR)-like lesion-inducer family protein, pollen allergen Lol p2 family protein, cysteine protease inhibitor, RNA polymerase subunit, and
Oryza sativa SKP1 (
S-phase kinase-associated protein 1)-like protein 1 (OSK1), so it is not clear whether the interactions are valid for the other proteins. Since the regulation network of these important protein-protein interactions is vital for the study of the function of OsUbc13 in depth, the follow-up work we are carrying out will be focused on confirming all the candidate
OsUbc13-interacting genes and then the function analysis of the re-tested genes where the function is unknown. In summary, our findings provide a preliminary foundation for understanding the function of OsUbc13.
Acknowledgements
We are grateful to Dr. Li-yu HUANG (Chinese Academy of Agricultural Sciences) for technical assistance and Dr. Sun-jie LU (Fudan University, Shanghai, China) for helpful suggestions in the discussion.
* Project supported by the National High-Tech R&D Program (863) of China (No. 2006AA10Z159) and the National Natural Science Foundation of China (No. 30871502) Ya WANG, Meng-yun XU, Jian-ping LIU, Mu-gui WANG, Hai-qing YIN, and Ju-min TU declare that they have no conflict of interest.References
[1] Agrawal, G.K., Abe, K., Yamazaki, M., 2005. Conservation of the E-function for floral organ identity in rice revealed by the analysis of tissue culture-induced loss-of-function mutants of the
OsMADS1 gene.
Plant Mol Biol, 59(1):125-135.


[2] Akashi, K., Nishimura, N., Ishida, Y., 2004. Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon.
Biochem Biophys Res Commun, 323(1):72-78.
[3] Al Bitar, F., Roosens, N., Smeyers, M., 2003. Sequence analysis, transcriptional and posttranscriptional regulation of the rice vdac family.
Biochem Biophys Acta, 1625(1):43-51.
[4] Andersen, P.L., Zhou, H., Pastushok, L., 2005. Distinct regulation of Ubc13 functions by the two ubiquitin-conjugating enzyme variants Mms2 and UEV1A.
J Cell Biol, 170(5):745-755.
[5] Apel, K., Hirt, H., 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction.
Annu Rev Plant Biol, 55(1):373-399.
[6] Ashley, C., Pastushok, L., McKenna, S., 2002. Roles of mouse UBC13 in DNA postreplication repair and Lys63-linked ubiquitination.
Gene, 285(1-2):183-191.
[7] Broomfield, S., Chow, B.L., Xiao, W., 1998.
MMS2, encoding a ubiquitin-conjugating-enzyme-like protein, is a member of the yeast error-free postreplication repair pathway.
PNAS, 95(10):5678-5683.
[8] Broomfield, S., Hryciw, T., Xiao, W., 2001. DNA postreplication repair and mutagenesis in
Saccharomyces cerevisiae
.
Mutat Res, 486(3):167-184.
[9] Brusky, J., Zhu, Y., Xiao, W., 2000.
UBC13, a DNA-damage-inducible gene, is a member of the error-free postreplication repair pathway in
Saccharomyces cerevisiae
.
Curr Genet, 37(3):168-174.
[10] Chau, V., Tobias, J.W., Bachmair, A., 1989. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein.
Science, 243(4898):1576-1583.
[11] Chen, Z.J., 2005. Ubiquitin signalling in the NF-κB pathway.
Nat Cell Biol, 7(8):758-765.
[12] Chen, Z.J., 2012. Ubiquitination in signaling to and activation of IKK.
Immunol Rev, 246(1):95-106.
[13] Chen, Z.J., Parent, L., Maniatis, T., 1996. Site-specific phosphorylation of IκBα by a novel ubiquitination-dependent protein kinase activity.
Cell, 84(6):853-862.
[14] Chien, C.T., Bartel, P.L., Sternglanz, R., 1991. The two-hybrid system: a method to identify and clone genes for proteins that interact with a protein of interest.
PNAS, 88(21):9578-9582.
[15] Dai, L., Xu, L., Huang, D., 2002. ASK1 physically interacts with COI1 and is required for male fertility in
Arabidopsis
.
Sci China C Life Sci, 45(6):631-636.
[16] Ehsani, A., Alluin, J.V., Rossi, J.J., 2013. Cell cycle abnormalities associated with differential perturbations of the human U5 snRNP associated U5-200kD RNA helicase.
PLoS ONE, 8(4):e62125
[17] Fields, S., Song, O., 1989. A novel genetic system to detect protein-protein interactions.
Nature, 340(6230):245-246.
[18] Finley, D., Ozkaynak, E., Varshavsky, A., 1987. The yeast poly-ubiquitin gene is essential for resistance to high temperatures, starvation and other stresses.
Cell, 48(6):1035-1046.
[19] Finley, D., Bartel, B., Varshavsky, A., 1989. The tails of ubiquitin precursors are ribosomal proteins whose fusion to ubiquitin facilitates ribosome biogenesis.
Nature, 338(6214):394-401.
[20] Galan, J.M., Haguenauer-Tsapis, R., 1997. Ubiquitin lys63 is involved in ubiquitination of a yeast plasma membrane protein.
EMBO J, 16(19):5847-5854.
[21] Geelen, D.N., Inz, D.G., 2001. A bright future for the bright yellow-2 cell culture.
Plant Physiol, 127(4):1375-1379.
[22] Godbole, A., Varghese, J., Sarin, A., 2003. VDAC is a conserved element of death pathways in plant and animal systems.
Biochim Biophys Acta, 1642(1-2):87-96.
[23] Godbole, A., Mitra, R., Dubey, A.K., 2011. Bacterial expression, purification and characterization of a rice voltage-dependent, anion-selective channel isoform, OsVDAC4.
J Membr Biol, 244(2):67-80.
[24] Grisvard, J., Aubusson-Fleury, A., Baroin-Tourancheau, A., 2010. Multiple uses of Lys63-polyubiquitination in the ciliate
Sterkiella histriomuscorum
.
Protist, 161(3):479-488.
[25] Guan, X., Diez, T., Prasad, T.K., 1999. Geranoyl-CoA carboxylase: a novel biotin-containing enzyme in plants.
Arch Biochem Biophys, 362(1):12-21.
[26] Hashimoto, M., Komatsu, K., Maejima, K., 2012. Identification of three MAPKKKs forming a linear signaling pathway leading to programmed cell death in
Nicotiana benthamiana
.
BMC Plant Biol, 12(1):103
[27] Hiraishi, H., Mochizuki, M., Takagi, H., 2006. Enhancement of stress tolerance in
Saccharomyces cerevisiae by overexpression of ubiquitin ligase Rsp5 and ubiquitin-conjugating enzymes.
Biosci Biotechnol Biochem, 70(11):2762-2765.
[28] Hochstrasser, M., 1996. Ubiquitin-dependent protein degradation.
Annu Rev Genet, 30(1):405-439.
[29] Hoege, C., Pfander, B., Moldovan, G.L., 2002. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO.
Nature, 419(6903):135-141.
[30] Hofmann, R.M., Pickart, C.M., 1999. Noncanonical MMS2-encoded ubiquitin-conjugating enzyme functions in assembly of novel polyubiquitin chains for DNA repair.
Cell, 96(5):645-653.
[31] Jentsch, S., 1992. The ubiquitin-conjugation system.
Annu Rev Genet, 26(1):179-207.
[32] Jentsch, S., McGrath, J.P., Varshavsky, A., 1987. The yeast DNA repair gene
RAD6 encodes a ubiquitin-conjugating enzyme.
Nature, 329(6135):131-134.
[33] Kim, J.A., Cho, K., Singh, R., 2009. Rice OsACDR1 (
Oryza sativa accelerated cell death and resistance 1) is a potential positive regulator of fungal disease resistance.
Mol Cells, 28(5):431-439.
[34] Lauber, J., Fabrizio, P., Teigelkamp, S., 1996. The HeLa 200 kDa U5 snRNP-specific protein and its homologue in
Saccharomyces cerevisiae are members of the DEXH-box protein family of putative RNA helicases.
EMBO J, 15(15):4001-4015.
[35] Lee, B.H., Kapoor, A., Zhu, J., 2006. STABILIZED1, a stress-upregulated nuclear protein, is required for pre-mRNA splicing, mRNA turnover, and stress tolerance in
Arabidopsis
.
Plant Cell, 18(7):1736-1749.
[36] Lee, L.Y., Fang, M.J., Kuang, L.Y., 2008. Vectors for multi-color bimolecular fluorescence complementation to investigate protein-protein interactions in living plant cells.
Plant Methods, 4(1):24
[37] Li, J., Wen, R., Andersen, P., 2010. Zebrafish Ubc13 is required for Lys63-linked polyubiquitination and DNA damage tolerance.
Mol Cell Biochem, 343(1-2):173-182.
[38] McKenna, S., Spyracopoulos, L., Moraes, T., 2001. Noncovalent interaction between ubiquitin and the human DNA repair protein Mms2 is required for Ubc13-mediated polyubiquitination.
J Biol Chem, 276(43):40120-40126.
[39] McKenna, S., Hu, J., Moraes, T., 2003. Energetics and specificity of interactions within Ub.Uev.Ubc13 human ubiquitin conjugation complexes.
Biochemistry, 42(26):7922-7930.
[40] Muralidhar, M.G., Thomas, J.B., 1993. The
Drosophila bendless gene encodes a neural protein related to ubiquitin-conjugating enzymes.
Neuron, 11(2):253-266.
[41] Murashige, T., Skoog, F., 1962. A revised medium for rapid growth and bioassays with tobacco cultures.
Physiol Plant, 15(3):473-493.
[42] Obata, K., Fukuda, T., Morishita, R., 2001. Human biotin-containing subunit of 3-methylcrotonyl-CoA carboxylase gene (MCCA): cDNA sequence, genomic organization, localization to chromosomal band 3q27, and expression.
Genomics, 72(2):145-152.
[43] Oh, C.E., McMahon, R., Benzer, S., 1994.
bendless, a
Drosophila gene affecting neuronal connectivity, encodes a ubiquitin-conjugating enzyme homolog.
J Neurosci, 14(5):3166-3179.
[44] Parusel, C.T., Kritikou, E.A., Hengartner, M.O., 2006. URI-1 is required for DNA stability in
C. elegans
.
Development, 133(4):621-629.
[45] Pastori, G.M., Foyer, C.H., 2002. Common components, networks, and pathways of cross-tolerance to stress. The central role of “redox” and abscisic acid-mediated controls.
Plant Physiol, 129(2):460-468.
[46] Pastushok, L., Xiao, W., 2004. DNA postreplication repair modulated by ubiquitination and sumoylation.
Adv Protein Chem, 69:279-306.
[47] Petersen, A., Suck, R., Lindner, B., 2006. Phl p 3: structural and immunological characterization of a major allergen of timothy grass pollen.
Clin Exp Allergy, 36(6):840-849.
[48] Pickart, C.M., 2001. Ubiquitin enters the new millennium.
Mol Cell, 8(3):499-504.
[49] Prasad, K., Parameswaran, S., Vijayraghavan, U., 2005. OsMADS1, a rice MADS-box factor, controls differentiation of specific cell types in the lemma and palea and is an early-acting regulator of inner floral organs.
Plant J, 43(6):915-928.
[50] Rothofsky, M.L., Lin, S.L., 1997. CROC-1 encodes a protein which mediates transcriptional activation from the human FOS promoter.
Gene, 195(2):141-149.
[51] Sato, S., Sanjo, H., Takeda, K., 2005. Essential function for the kinase TAK1 in innate and adaptive immune responses.
Nat Immunol, 6(11):1087-1095.
[52] Schweimer, K., Petersen, A., Suck, R., 2008. Solution structure of Phl p 3, a major allergen from timothy grass pollen.
Biol Chem, 389(7):919-923.
[53] Sheen, J., 2001. Signal transduction in maize and
Arabidopsis mesophyll protoplasts.
Plant Physiol, 127(4):1466-1475.
[54] Silva, A.T., Nguyen, A., Ye, C., 2010. Conjugated polymer nanoparticles for effective siRNA delivery to tobacco BY-2 protoplasts.
BMC Plant Biol, 10(1):291
[55] Song, J., Wurtele, E.S., Nikolau, B.J., 1994. Molecular cloning and characterization of the cDNA coding for the biotin-containing subunit of 3-methylcrotonoyl-CoA carboxylase: identification of the biotin carboxylase and biotin-carrier domains.
PNAS, 91(13):5779-5783.
[56] Steller, H., 1995. Mechanisms and genes of cellular suicide.
Science, 267(5203):1445-1449.
[57] Susin, S.A., Zamzami, N., Castedo, M., 1997. The central executioner of apoptosis: multiple connections between protease activation and mitochondria in Fas/APO-1/CD95- and ceramide-induced apoptosis.
J Exp Med, 186(1):25-37.
[58] Takacs, E.M., Suzuki, M., Scanlon, M.J., 2012. Discolored1 (DSC1) is an ADP-ribosylation factor-GTPase activating protein required to maintain differentiation of maize kernel structures.
Front Plant Sci, 3:115
[59] Tamura, K., Dudley, J., Nei, M., 2007. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0.
Mol Biol Evol, 24(8):1596-1599.
[60] Tokunaga, F., Sakata, S., Saeki, Y., 2009. Involvement of linear polyubiquitylation of NEMO in NF-κB activation.
Nat Cell Biol, 11(2):123-132.
[61] Tripathi, P., Rabara, R.C., Lin, J., 2013. GmWRKY53, a water- and salt-inducible soybean gene for rapid dissection of regulatory elements in BY-2 cell culture.
Plant Signal Behav, 8(5):e24097
[62] Ulrich, H.D., Jentsch, S., 2000. Two RING finger proteins mediate cooperation between ubiquitin-conjugating enzymes in DNA repair.
EMBO J, 19(13):3388-3397.
[63] Wang, W.M., Ma, X.F., Zhang, Y., 2012. PAPP2C interacts with the atypical disease resistance protein RPW8.2 and negatively regulates salicylic acid-dependent defense responses in
Arabidopsis
.
Mol Plant, 5(5):1125-1137.
[64] Wang, Y., Zhang, X.B., Lu, S.J., 2012. Inhibition of a basal transcription factor 3-like gene Osj10gBTF3 in rice results in significant plant miniaturization and typical pollen abortion.
Plant Cell Physiol, 53(12):2073-2089.
[65] Weaver, L.M., Lebrun, L., Franklin, A., 1995. Molecular cloning of the biotinylated subunit of 3-methylcrotonyl-coenzyme A carboxylase of
Arabidopsis thaliana
.
Plant Physiol, 107(3):1013-1014.
[66] Wei, W., Ayad, N.G., Wan, Y., 2004. Degradation of the SCF component Skp2 in cell-cycle phase G1 by the anaphase-promoting complex.
Nature, 428(6979):194-198.
[67] Weil, R., Israel, A., 2004. T-cell-receptor and B-cell-receptor-mediated activation of NF-κB in lymphocytes.
Curr Opin Immunol, 16(3):374-381.
[68] Wen, R., Newton, L., Li, G., 2006.
Arabidopsis thaliana UBC13: implication of error-free DNA damage tolerance and Lys-63-linked polyubiquitylation in plants.
Plant Mol Biol, 61(1-2):241-253.
[69] Wen, R., Torres-Acosta, J.A., Pastushok, L., 2008.
Arabidopsis UEV1D promotes Lysine-63-linked polyubiquitination and is involved in DNA damage response.
Plant Cell, 20(1):213-227.
[70] Wong, H.L., Sakamoto, T., Kawasaki, T., 2004. Down-regulation of metallothionein, a reactive oxygen scavenger, by the small GTPase OsRac1 in rice.
Plant Physiol, 135(3):1447-1456.
[71] Wu, X., Yamamoto, M., Akira, S., 2009. Regulation of hematopoiesis by the K63-specific ubiquitin-conjugating enzyme Ubc13.
PNAS, 106(49):20836-20841.
[72] Xiao, W., Lin, S.L., Broomfield, S., 1998. The products of the yeast MMS2 and two human homologs (hMMS2 and CROC-1) define a structurally and functionally conserved Ubc-like protein family.
Nucl Acids Res, 26(17):3908-3914.
[73] Yamaguchi, T., Kim, N.S., Sekine, S., 1996. Cloning and expression of cDNA encoding a human ubiquitin-conjugating enzyme similar to the
Drosophila bendless gene product.
J Biochem, 120(3):494-497.
[74] Yamamoto, M., Sato, S., Saitoh, T., 2006. Cutting edge: pivotal function of Ubc13 in thymocyte TCR signaling.
J Immunol, 177(11):7520-7524.
[75] Yamamoto, M., Okamoto, T., Takeda, K., 2006. Key function for the Ubc13 E2 ubiquitin-conjugating enzyme in immune receptor signaling.
Nat Immunol, 7(9):962-970.
[76] Yang, R.F., Tang, Q.C., Wang, H.M., 2011. Analyses of two rice (
Oryza sativa) cyclin-dependent kinase inhibitors and effects of transgenic expression of OsiICK6 on plant growth and development.
Ann Bot, 107(7):1087-1101.
[77] Yang, Y., Liu, Z.H., Ware, C.F., 1997. A cysteine protease inhibitor prevents activation-induced T-cell apoptosis and death of peripheral blood cells from human immunodeficiency virus-infected individuals by inhibiting upregulation of Fas ligand.
Blood, 89(2):550-557.
[78] Yang, Z., Wu, Y., Li, Y., 2009. OsMT1a, a type 1 metallothionein, plays the pivotal role in zinc homeostasis and drought tolerance in rice.
Plant Mol Biol, 70(1-2):219-229.
[79] Yoshida, S., Forno, D.A., Cock, J.H., 1976. Routine procedures for growing rice plants in culture solution. Laboratory Manual for Physiological Studies of Rice. IRRI,Los Baños, Philippines :61-66.
[80] Yuan, J., Chen, D., Ren, Y., 2008. Characteristic and expression analysis of a metallothionein gene,
OsMT2b, down-regulated by cytokinin suggests functions in root development and seed embryo germination of rice.
Plant Physiol, 146(4):1637-1650.
[81] Yuan, S., Fu, Y., Wang, X., 2008. Voltage-dependent anion channel 1 is involved in endostatin-induced endothelial cell apoptosis.
FASEB J, 22(8):2809-2820.
[82] Zang, Y., Wang, Q., Xue, C., 2012. Rice
UBC13, a candidate housekeeping gene, is required for K63-linked polyubiquitination and tolerance to DNA damage.
Rice, 5(1):24-34.



Open peer comments: Debate/Discuss/Question/Opinion
<1>